Well, if he is to be believed, it would seem the case that in the first instance, he thinks all relevantly-interested biologists are wrong simply because he is obsessively convinced his personal origin narrative must be correct, with the moronic religion-based rationalisations following afterwards.
Those moronic rationalisations including:
His narrative being especially compatible with a Deity/Nature Spirit, even though essentially any other narrative is just as amenable to 'godwashing' as his, especially given how vague his deity-as-force-of-nature pseudogod is.
His narrative being overlooked by essentially every biologist for the last century because despite supposedly being obvious, it wasn't sufficiently 'self-creative', despite the fact that by any reasonable reading it is at least as 'self-creative' as the theories he complains about.
Indeed, if factoring in his hopeless wibbling about our ancient ancestors habitually training with weapons largely or solely in the expectation of occasional future predator encounters, and changing their all-important dentition as a result, it seems rather more 'self-creative' than most.
How atheist ideology messed up the human origin story
Spin-off from "Dialog on 'Creationists read this' "
Moderators: kiore, Blip, The_Metatron
Re: How atheist ideology messed up the human origin story
#1001 ![]() by tolman » Dec 10, 2015 1:04 pm
by tolman » Dec 10, 2015 1:04 pm
I don't do sarcasm smileys, but someone as bright as you has probably figured that out already.
- tolman
- Posts: 7106
- Country: UK
Re: How atheist ideology messed up the human origin story
#1002 ![]() by Animavore » Dec 10, 2015 2:51 pm
by Animavore » Dec 10, 2015 2:51 pm
THWOTH wrote:Animavore wrote:I'm sorry. I haven't looked up this thread in ages. What the fuck is going on? What does any of this small/large canine stuff have to do with creationism? In a nutshell.
Currently jayjay#'s is maintaining that the dentition of proto-humans and contemporary hominids demonstrates that the scientific consensus about our species evolutionary development and lineage is errant, having been unduly influenced by a wilful and short-sighted desire to exclude the deity of a particular tribe of iron age, Middle-Eastern goat bothers as a possible, and indeed by his lights more probable, explanation for both our species existence and all life on Earth.


A most evolved electron.
-

Animavore - Name: The Scribbler
- Posts: 45108
- Age: 45

")
Re: How atheist ideology messed up the human origin story
#1003 ![]() by Darwinsbulldog » Dec 10, 2015 3:18 pm
by Darwinsbulldog » Dec 10, 2015 3:18 pm
THWOTH wrote:Animavore wrote:I'm sorry. I haven't looked up this thread in ages. What the fuck is going on? What does any of this small/large canine stuff have to do with creationism? In a nutshell.
Currently jayjay#'s is maintaining that the dentition of proto-humans and contemporary hominids demonstrates that the scientific consensus about our species evolutionary development and lineage is errant, having been unduly influenced by a wilful and short-sighted desire to exclude the deity of a particular tribe of iron age, Middle-Eastern goat bothers as a possible, and indeed by his lights more probable, explanation for both our species existence and all life on Earth.
Well, those goat-herders [or similar group] did evolve adult lactose tolerance, so I suppose they were the origin of yet more evolutionary pie to throw in the creationist's faces!
Jayjay4547 wrote:
"When an animal carries a “branch” around as a defensive weapon, that branch is under natural selection".
"When an animal carries a “branch” around as a defensive weapon, that branch is under natural selection".
- Darwinsbulldog
- Posts: 7440
- Age: 69
Re: How atheist ideology messed up the human origin story
#1004 ![]() by Jayjay4547 » Dec 14, 2015 4:45 am
by Jayjay4547 » Dec 14, 2015 4:45 am
Calilasseia wrote:Oh look, it's Lies And Bullshit TimeTM again, brought to your TV by your regular host JayJay ...
After an opening like that, I don’t expect a civil or reasonable discussion from the rest of your post.
Calilasseia wrote:Jayjay4547 wrote:But my point is(a) that the lack of fangs in Australopithecus should have immediately created the inference that they instead used hand held weapons that made them dangerous to attack.
Bollocks. First of all, there are plenty of organisms that don't possess big canines, and which don't use tools.
Armadillos would be an example of animals that don’t possess big canines and don’t use tools. The issue isn’t about “tools” in general, but specifically about hand held weapons, used to make a species dangerous to attack. For that, an animal would need hands capable of grasping a weapon, means to carry a weapon around, it would need to be of a certain absolute size where kinetic weaponry can be effective, and a certain size relative to predator. Australopithecus satisfied those criteria, and higher primates generally are near the edge of it, shown by Kortlandt’s 1960s experiments with chimps and a dummy leopard, and by Boesch’s account of chimps taunting a cornered leopard using sticks. We have been over those a few times. Anyway I came across a cute supporting anecdote the other day, about someone who, hoping to protect his car bonnet and windscreen wipers from damage by baboons in the Cape Point nature reserve, left a rubber snake on the dashboard. When he got back to his car he found the windscreen had been smashed by baboons, using stones.
Calilasseia wrote: Second, Sahelanthropus had small canines, but never once during its 3 million year history left behind evidence of tool use. Oh dear, there goes your fantasy once again, destroyed by DATA.
I will get back to Sahelanthropus canines a bit later but really its no more needed to find tools tied to an ancestor of Australopithecs, than one needs to find bones stuck in the teeth of T.rex to figure out it ate meat. Palaeontologists had not trouble figuring that out, long before they found marks left by those animals. On top of that, Your “3 million year history” is more correctly a 3 million year GAP between the date of one fossil actual or near ancestor, and Australopithecus. You demand for evidence, a tool found in that gap, to be positively tied to Sahelanthropus, for lack of which you pronounce my “fantasy” destroyed by “DATA”.
Calilasseia wrote:Jayjay4547 wrote: It’s nonetheless true that the long sharp canines of other higher primates do make them dangerous attack. They are ferocious biters.
Oh wait, how often have you been presented with DATA, JayJay, demonstrating that a significant proportion of the use of those canines, focuses on competition between rivals of the same species for a highly desirable resource?
Indeed, how many primate species exhibit manifest sexual dimorphism in this regard, which makes NO sense whatsoever in the light of your obsessive concentration on defensive biting as purportedly the sole reason for this dentition, even before we factor in such things as diet? Because one of the elementary thoughts that immediately crosses the mind of anyone who has paid attention in biology class, is that a defensive capability of this sort would be useful to members of a given species regardless of gender, if a pressing need accompanied by a measurable selection pressure was present for said capability. The mere fact that something as functional as dentition, undergoes gender biased modification, tells those of us who paid attention in biology class, that your obsessive fantasising about defensive biting is precisely that, because modification of this sort would not affect significantly, a feature already subject to an alternative strong selection pressure.
To repeat, I’ve sidestepped that whole trope about sexual selection by just sticking to the point that the long sharp canines of other higher primates make them dangerous to attack, which is significant for their ecology.. Consider that video clip I showed about humans very circumspectly trying to attack a group of baboons occupying and then rifling a car. The large male on top of the car intimidated the men, keeping them away until the baboons had got what they wanted. The observation by Cheney and others that all the members of troop get involved in a mobbing attack doesn’t contradict that the sexually dimorphous male primates are the ones who can damage- and everyone knows that instinctively.
Here is a problem for you. Suppose that primate sexual dimorphism in canines is driven by sexual selection. Why is it then that men and ancestral Australopithecus males lack long canines? Why is it that we men have inherited such girly skulls? Were Australopithecus utterly above such crude competition for the possession of females, as Darwin put it? Seems to me that the Australopithecus males competed using stick fights just like young tribal men today. And those skills were turned against predators, making a hominin troop dangerous to attack. Intraspecific competition in primates and access to females is just a cruel trick by “nature” to keep testosterone filled individuals from breeding out of the gene pool.
Calilasseia wrote: But this is another piece of DATA you keep ignoring whilst trying to peddle your sad little fantasy as purportedly constituting fact,
It’s not a fantasy, not sad, or even little. It’s a strong INFERENCE that Australopithecus used hand held weapons to make them dangerous to attack. You used that word “fantasy” 16 times in your post.
Calilasseia wrote: regardless of how much DATA destroys that fantasy. DATA such as:
1: NO hominid weapons older than 100,000 years have ever been found;
When I look at your so called DATA it turns out to date the appearance of hafted weapons used for hunting by fully modern Homo sapiens. That’s only a bit less ridiculous than demanding to see a rifle before admitting weaponry.
Calilasseia wrote: 2: ALL tools reliably associated with Australopithecus have been FOOD PREPARATION TOOLS;
One of the uses of an Oldowan hand axe would have been to ding the skull of a predator. Edged tools have been used for woodworking as you say later in your post- that isn’t food preparation. What do you imagine Australopithecus might have whittled away at wood FOR? The most plausible task would have been, to fashion a pointed stick to keep predators at arms length. Sure as hang, if you slept in a cave and a leopard appeared at the entrance, who had already eaten your aunt and two nephews, and you could whittle away at wood, you would make a pointy stick.
Calilasseia wrote:Jayjay4547 wrote:The other day I asked a local expert on primates “Do primates bite?” and she said yes, and that her body is full of scars as evidence of that. When I looked tremendously interested she thought it was her body that excited my interest and became a bit stiffly cold.
If your approaches to her were the same as your approaches to us with respect to the proper conduct of discourse, I'm not in the least surprised about that last part.
My approaches to posters here doesn’t include calling anyone+- a liar, a masturbatory fantasist, talking bullshit, etc etc. Your concept of “proper conduct of discourse” involves zero commitment to civility. Besides that, you sidestep that expert’s account did support the view that other primates are proficient and ready biters.
.Calilasseia wrote:Jayjay4547 wrote:Here is a pic of Sahelanthropus showing that it had longer canines than Australopithecus , suggesting that the adaptation was in progress roughly around that time.
A New Hominid From The Upper Miocene Of Chad, Central Africa by Michel Brunet, and others here]Brunet et al, 2002 wrote:[b]Differential diagnosis. Sahelanthropus is distinct from all living great apes in the following respects: relatively smaller canines with apical wear, the lower showing a full occlusion above the well-developed distal tubercle, probably correlated with a non-honing C–P3 complex (P3 still unknown).Brunet et al, 2002 wrote:Sahelanthropus is distinguished as a hominid from large living and known fossil hominoid genera in the following respects: …from Gorilla by smaller canines and lower-cusped cheek teeth
]from Ouranopithecus…by markedly smaller but mesiodistally longer canines, apical wear and large distal tubercle in lower canines
Frim Sivapithecus16, smaller canines with apical wear, and thinner cheek-teeth enamel
Sahelanthropus is also distinct from all known hominid genera in the following respects:
from Homo ….non-incisiform and mesiodistally long canines,
from Ardipithecus[/i….; less incisiform upper canines not diamond shaped with a low distal shoulder and a mesiodistal long axis, bucco-lingually narrower lower canines with stronger distal tubercle, and P4 with two roots;….
from [i]Orrorin … non-chimp-like upper canines with extensive apical wear.
Again, note the frequent references to small canines with apical wear

I extracted from your quote above, the bits directly relating to canines. They do support my claim that this possible ancestors of Australopithecus had canines intermediate between those of apes and Australopithecus. Here’s a pic of another possible ancestor Ardipithecus Ramidus- without canines that show “extensive apical wear” (ie worn down from on top)
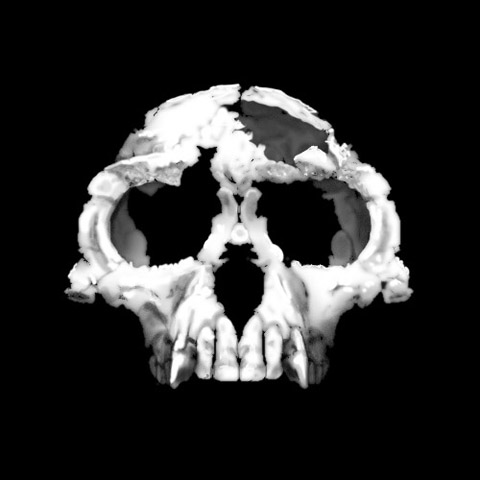
It’s not essential to pin down the habits of ancestors of Australopithecus, to see that even the early Australopithecus afarensis canines shows that they had already fully responded to relief from having to bite their predators.
Calilasseia wrote:Shea, 1988 wrote:Lithic use-wear analysis can provide functional infomation about stone tool assemblages and insights into prehistoric hominid behaviour. The criteria by which hafter stone spear- and arrow-points can be recognised have been extensively documented by other researchers (Ahler 1971; Fischer, Hansen and Rasmussen 1984; Odell and Cowan 1986). Indentifying such wear patterns in the Middle Palaeolithic can indicate both the geographic and temporal variation of prehistoric hunting strategies. Wear patterns on stone tools from the Levantine Middle Palaeolithic sites of Kebara, Qazfeh, and Hayonim caves suggest that technologically-assisted hunting may have been an important part of hominid subsistence as early as 50,000-100,000 years ago.
Oh wait, what did I spend several posts in this thread schooling you on? LITHIC WEAR ANALYSIS. Namely, the analysis of the different tool usage marks that different tool usages leave upon the tools in question, and which scientists have been pressing into service to analyse ACTUAL tool usage. Yet more DATA you keep ignoring. Indeed, I even schooled you on the fact that scientists went to the trouble of making their own prehistoric stone tools, in accordance with the known toolmaking methods deployed by earlier hominids, and then subjected those tools to a range of usages, to provide a database of use-wear analysis data. Indeed, I told you about this in this same post in which I provided the paper discussing the use of this technique to determine actual tool usage. I cited no less than four peer reviewed scientific works covering this topic in that post, and presented the full papers in two of those instances. So your assertion that I didn't back up my statement is a BARE FACED LIE.
This is all you have to offer here, JayJay, made up shit accompanied by bare faced lies and other well-documented forms of creationist duplicity. I and others here, on the other hand, have REAL DATA to bring to the table.
Now, since I've already made numerous references to lithic wear analysis, and how this allows scientists to distinguish which usage a given tool was put to, I think we can all safely conclude that I've provided more than enough substance allowing a weapon to be distinguished from, say, a food preparation or materials handling tool, because, lo and behold, as scientists have discovered experimentally, all of these usages leave distinct and analysable wear patterns on the tools in question. A weapon would have different usage marks to a food preparation tool, which in turn would have different usage marks to a materials processing tool. Indeed, the Shea paper I provided earlier mentions a specific experiment conducted to establish use-wear patterns on stone tools, viz:Shea, 1988 wrote:Whatver approach or combination of approaches is employed, archaeological analyses must be accompanied by a demonstration of each analyst's ability to reliably reconstruct the functions of experimentally-utilised stone tools. It is only from such a demonstration that accuracy levels and confidence intervals can be assigned to archaeological interpretations. In the course of learning the "low magnification" approach, six independent blind tests of the author's ability to reconstruct stone tool uses were conducted over three years (Shea 1987). These tests involved a total of 243 uses on 111 artefacts (with 33 unused "dummy" specimens). All artefacts had been used, without the author's participation or observation, by graduates of lithic technology workshops and had been subjected to simulated archaeological screening and washing. The test results suggest the following accuracy levels and confidence intervals for archaeological application: location of use = 237/243 correct (x=97%±1%); action employed (of 16 possibilities) = 222/243 correct (x=91%±5%), worked material (of 12 possibilities) = 200/243 correct (x=82%±5%). No statistically significant differences were noted in the accuracy rates for different actions and worked materials, or between retouched and unretouched edges. Only the duration of tool use and the type of lithic raw material appeared to significantly affect analytical accuracy.
So the author not only engaged in an experiment aimed at determining which wear patterns accompany which tool usage, but also aimed at determining his ability to deduce correctly the requisite wear usage. Now the mere fact that I brought that paper here, on its own tells anyone appraising this thread honestly, that I did more than sufficient to establish my case. Namely, that scientists have rigorous methods allowing them to tell the difference between tools used as weapons, tools used for food preparation, and tools used for material processing by prehistoric hominids. They have the DATA. The mere fact that you couldn't be bothered to examine that paper, alight upon this DATA yourself, then fabricated a bare faced lie to the effect that I failed to support my statements robustly in this vein, again destroys your fantasy.
I did miss your citing a paper that dated the earliest weapons to 100 000 years ago, being hafted hunting weapons. As to lithic wear patterns, a lithic laboratory workshop in which participants didn’t face predation would be a markedly poor reconstruction of Australopithecus natural context. So the possible use of an Oldowan hand axe to smash a predator skull wasn’t even considered. And if a hand axe was used for that purpose, it might well not have been used often enough to show wear patterns,
Calilasseia wrote:
Which, funnily enough, is followed shortly after by the entire section on evidence from wear analysis that you never bothered reading. I wonder why you never bothered reading it? would this have something to do with the fact that all the scientific literature pertinent to this DESTROYS your fantasy assertions?

The only fantasy around here is your overbearing fantasy about destroying propositions that you don’t like.
Calilasseia wrote:Jayjay4547 wrote:Would a primate holding a hand axe like that, be dangerous to attack? My sense is yes but with a major reservation; something is missing; a means to keep a predator at arms length.
Oh wait, what usage wear analysis exists to support your fantasy about weapon wielding Australopithecines? NONE.
A stone like that could have crushed many a skull without showing wear patterns. Answer the question. Or let me put it another way: Can you imagine a better shape for a stone intended and held so as to crush a skull?
Calilasseia wrote:Jayjay4547 wrote:That primate would need a stopper tool either in its other hand or held in two hands by another member of the troop. Think of a dog attacking you- you would need something to stop it with and something to punish it with. But of course, sticks unlike stones are highly perishable and relatively unlikely to be found as fossils.
HA HA HA HA HA HA HA!
JayJay, we have in existence FOSSILS OF TREES DATING BACK ALL THE WAY TO THE CARBONIFEROUS. Under the right conditions, wood can fossilise, and persist for OVER 300 MILLION YEARS. The oldest known tree, Watteiza, dates back all the way to the middle Devonian, 385 million years before present. Archaeopteris is only slightly younger at 383 million years.
If you want to play that game, the oldest stones date back nearly ten times longer than your oldest fossil wood. Oldowan hand axes are preserved till today except for a chance finding their way into a grinding stream bed or glacial floor. But whatever sticks Australopithecus made using stone tools, would only be preserved under exceptional circumstances.
Calilasseia wrote: But of course, none of this in the least supports your sad little fantasy, about Australopithecines wielding weapons like some sort of prehistoric Call of Duty spec-ops troop. All the DATA says your fantasy never happened.
Call of Duty? That might be your fantasy, but I haven’t presented Australopithecus that way. A weird habit of Australopithecus enabled their males to have girly skulls, and so unlike other primates, freed them from holding up evolution of the species. OK that’s just a shadow of the truth but it’s a lot more true than the story line in Darwin’s Descent of Man where female allure provoked their menfolk into evolving.
-

Jayjay4547 - THREAD STARTER
- Name: Jonathan
- Posts: 1482
- Country: South Africa
")
Re: How atheist ideology messed up the human origin story
#1005 ![]() by Jayjay4547 » Dec 14, 2015 4:57 am
by Jayjay4547 » Dec 14, 2015 4:57 am
Fenrir wrote:I'm left handed.
Does that mean i have an offensive defense?
Or a defensive offense?
:shrug
I'm also left handed. It could mean that one is a bit more sinister,(sinistrous) tricky, defensive minded, lacking in killer instinct. Or not.
-
Jayjay4547 - THREAD STARTER
- Name: Jonathan
- Posts: 1482
- Country: South Africa
Re: How atheist ideology messed up the human origin story
#1006 ![]() by Thomas Eshuis » Dec 14, 2015 5:28 am
by Thomas Eshuis » Dec 14, 2015 5:28 am
Whinging about tone won't hide your disingenuous dodging Jayjay.
If you did not consistently present a bigoted and dishonest narrative, you wouldn't get treated as a person who consistently presents a dishonest and bigoted narrative.
You don't get to demand or expect civility or reasonable discussion when you yourself present anything but.
"Respect for personal beliefs = "I am going to tell you all what I think of YOU, but don't dare retort and tell what you think of ME because...it's my personal belief". Hmm. A bully's charter and no mistake."
-

Thomas Eshuis - Name: Thomas Eshuis
- Posts: 31091
- Age: 34
- Country: Netherlands
")
Re: How atheist ideology messed up the human origin story
#1007 ![]() by Sendraks » Dec 14, 2015 10:39 am
by Sendraks » Dec 14, 2015 10:39 am
Jayjay4547 wrote:
It’s not a fantasy, not sad, or even little. It’s a strong INFERENCE that Australopithecus used hand held weapons to make them dangerous to attack. You used that word “fantasy” 16 times in your post.
Changing the word "fantasy" to "inference" doesn't make your argument any stronger JayJay. Whichever word you use, you're still making stuff up in an evidence free environment.
Jayjay4547 wrote: So the possible use of an Oldowan hand axe to smash a predator skull wasn’t even considered. And if a hand axe was used for that purpose, it might well not have been used often enough to show wear patterns,
Ahhh, classic theist argument. "just because you can't see the evidence doesn't mean it didn't happen/doesn't exist/isn't there."
My inference is that Australopiths defended themselves using a form of highly sophisticated hand to hand combat, which employed their great strength to good effect. This combat form would've been, at least, as formidable as anything developed in China millions of years later.
Of course the bones of Australopiths don't show any signs of these formidable technique being used to deter predators, but that probably only happened rarely, which explains that away and alleviates me from the burden of evidence.
It also alleviates me from the burden of rational discourse or the burden of anyone taking my views seriously.
I feel wonderously floaty and light free of these constraints.
Like myself and my arguments suddenly lack all substance.
"One of the great tragedies of mankind is that morality has been hijacked by religion." - Arthur C Clarke
"'Science doesn't know everything' - Well science knows it doesn't know everything, otherwise it'd stop" - Dara O'Brian
"'Science doesn't know everything' - Well science knows it doesn't know everything, otherwise it'd stop" - Dara O'Brian
-

Sendraks - Name: D-Money Jr
- Posts: 15260
- Age: 107
- Country: England
Re: How atheist ideology messed up the human origin story
#1008 ![]() by THWOTH » Dec 14, 2015 8:11 pm
by THWOTH » Dec 14, 2015 8:11 pm
Your Woo-Fu is strong master Sendraks, your logic formidable. 
"No-one is exempt from speaking nonsense – the only misfortune is to do it solemnly."
Michel de Montaigne, Essais, 1580
Michel de Montaigne, Essais, 1580
-

THWOTH - RS Donator
- Posts: 38753
- Age: 59
- Country: Untied Kingdom
")
Re: How atheist ideology messed up the human origin story
#1009 ![]() by Calilasseia » Dec 15, 2015 2:43 pm
by Calilasseia » Dec 15, 2015 2:43 pm
Oh this is going to be fun ...
As this is a long post, it's a two parter. Welcome to Part 1.
Well given that much of your output has repeatedly been demonstrated to be lies and bullshit, courtesy of the DATA telling us this, I'm merely taking this fact on board in advance before going on to demolish more of the lies and bullshit you post. Much of which turned out to be lies and bullshit once again once I addressed it. In short, JayJay, if you don't want your output to be regarded as duplicitous and worthy only of scorn and derision as a result, then there's a simple answer - instead of engaging in synthetic whingeing and bleating about post style when you're caught in flagrante delicto, stop posting manifest falsehoods and trying to pass them off as fact. Or is this simple answer too difficult for you even to acknowledge the existence of, let alone try to implement?
Oh, and as for a "reasonable" discussion, JayJay, my idea of a "reasonable" discussion, is one in which proper evidence is presented to support ideas, not made up shit. By that criterion, your entire posting career here has been wholly unreasonable, yet, instead of merely whingeing and bleating about this, I and the others here have exercised the diligent effort to demonstrate that your posts are lies and bullshit from start to finish. This is how discourse is conducted by grown ups.
Oh I can think of plenty of others. How about this for a nice selection?
Starting with the placental mammals, we'll run with, say, Heterohyrax brucei, shall we?

Or how about Rhyncocyon petersi, the Black & Rufous Elephant Shrew?

Or Galeopterus variegatus, the Colugo or Sunda Flying Lemur?
,_Central_Catchment_Area,_Singapore_-_20060618.jpg)
Or how about the Dugong, Dugong dugon?

Or for that matter, its close relative, the Antillean Manatee, Trichechus manatus?

Oh, but we forgot the marsupials! Have a Koala Phascolarctos cinereus, for example:

Or the Sugar Glider, Petaurus breviceps:

Or Antechinus stuarti, one of several marsupial mice in the Genus, famous for its males literally shagging themselves to death when reaching sexual maturity:

Or the unique Dromiciops gliroides, the sole living representative of the Microbiotheria (which also has no less than 50 teeth):

Then we have the Platypus, Onithorhynchus anatinus, a unique still-extant Monotreme :

What about other Vertebrates? Try this one - a female Black-Chinned Hummingbird, Archilochus alexandri ... there are 338 species of hummingbird known to science currently:

Or the Red-Crested Turaco, Tauraco erythrolophus:

Or the Bar-Throated Minla, Minla stigula:

Or perhaps, moving on to reptiles, we could choose this one, Cercosaura argulus :

Here's another nice organism lacking canines or tool usage. Say hello to Forcipiger longirostris from Hawaii:

Or how about this one? Say hello to Corydoras panda, which apart from being a species I've bred in the aquarium, also lacks oral dentition of any sort:

Perhaps a better candidate would be Beaufortia kweichowensis, one of the hillstream loaches, which again lacks oral dentition of any sort, and for that matter lacks any form of armour, being a scaleless fish? Here it is:

Just for the record, the Cobitidae, Balitoridae, Nemacheilidae and Gastromyzontidae, are all scaleless fishes. As are many of the catfishes in the Order Siluriformes. In the case of the loaches, the taxonomy is somewhat in flux at the moment, while Kottelat performs his major revision of the entire clade. But oops, we left out the INvertebrates! Let's remedy this shall we?
Well start with Carausius morosus, the Indian Stick Insect:

Or how about Cethosia nietneri?

(I won't bother with any more insects, as I have something like 40,000 photos of these on my hard drive as it is ... )
)
Here's another invertebrate, in the form of Anadenobolus monilicornis :

Then there's this one, Daphnia pulex :

Or perhaps Argulus foliaceus is more to your liking?

Or how about Chromodoris annae, one of many species of nudibranch?

Or Elysia crispata, if you prefer something a little more rococo in appearance?

Or Pteraeolidia ianthina?

Or try Schmidtea mediterranea, which is being added to the list of laboratory model organisms by genetics researchers ...

Or how about Caenorhabditis elegans, which has been so informative to scientists in recent decades?

Finally, how about these sponges?

I was going to toss into the mix this beast, namely one of the Warnowiids:

but this takes us outside the Metazoa, because dinoflagellates are algae, and therefore plants, though in the case of Warnowiids, they're not only mobile but have eyes. Just to illustrate how diverse the biosphere really is, and that viewing its contents in the same manner as a three year old's pop-up book on Noah's Ark will merely elicit much pointing and laughing. Which provides much of the reason for me carpet bombing you with all those photos, because all too often, your view of the biosphere is that of a three-year-old's pop up book on Noah's Ark. Though perhaps not as floridly so as American creationists, whose thinking about the biosphere consists of thinking that the rest of the species on the planet were put here to provide them with burgers and fries. But a timely reminder that we're not so fucking special after all is apposite here.
Moving on ...
Except that once again, NO EVIDENCE OF SUCH WEAPONS OLDER THAN 100,000 YEARS BEFORE PRESENT HAS EVER BEEN FOUND. Game. Fucking. Over.
It would also need to be capable of formulating the idea. Except that oops, we have NO EVIDENCE that hominids of any taxonomic designation, formulated the idea of developing hand held weapons until just 100,000 years ago. Game. Fucking. Over.
Except that, oops, NO EVIDENCE OF HAND HELD WEAPONS FASHIONED BY AUSTRALOPITHECINES EXISTS. Game. Fucking. Over.
Which I've already questioned ruthlessly when you first posted this. Funny how you never bothered to answer this with substance.
Oh wait, what do all of these instances have in common? Social organisation. Which the DATA tells us appears to be required to be in place for a protracted period of time first.
And I demolished your made up shit then as well.
Know this for a fact, does he? Was this actually recorded happening?
Oh, you mean you're going to make more shit up on this subject, despite me having presented the original scientific paper describing the species, in which the canines were determined to be no bigger than 11 mm? Oh that's going to be fucking hilarious when you do.
Except that, oh wait, scientists didn't just fucking ASSUME that T.rex teeth were indicative of a carnivorous trophic niche, they set about acquiring DATA to establish this. Starting with a hell of a lot of time spent making comparative anatomy a rigorous discipline. Then, lo and behold, they also found fossils of prey organisms bearing bone marks consistent with T. rex attack.
Oh wait, what were those two words I posted above again? Comparative anatomy. Which had been in place for some 100 years or so before palaeontology really took off. Which in turn became possible thanks to Linnaeus launching the taxonomic enterprise, and encouraging concentration on detail.
Er, wrong. Apart from the fact that gaps are a known creationist fetish, we've plenty of evidence for species persisting for far longer periods than 3 million years, and indeed, 3 million years would be an entirely reasonable time scale for a transition from Sahelanthropus to Australopithecus. You're also forgetting Ardipithecus, another pre-Australopithecine hominid Genus, with A. ramidus dating to 4.4 million years before present, and A. kadabba, dating to 5.6 million years before present. None of these organisms exhibited any propensity for tool use either.
It's time for another scientific paper, namely:
Re-Examining Human Origins In Light Of Ardipithecus ramidus by C. Owen Lovejoy, Science, 326: 74-74e8 (2nd October 2009) [Full paper downloadable from here]
From that paper in more detail:
The author provides this in the preamble article preceding the paper:
Moving back to the paper itself, we have this:
From the diagram on page 74e4, we have this:
The paper continues as follows:
So the author presents the dynamics of reproduction within a social group as being primary determinants of the requisite features, not your fantasy acquisition of weapons, and presents data supporting this linkage.
From the conclusion:
Looks like once again, your assertions have received a thorough fisking from real science.
Oh wait, what happened when tools were found that could be reliably associated with Australopithecus? Analysis of those tools revealed them to be food preparation tools. For a long time, even these were unknown to be associated with this taxon. But given the ubiquity of feeding related tools throughout the Eumetazoa, it's not surprising to those of us who paid attention in biology class, that the first tools to be found in association with Australopithecus should turn out to be tools of this sort. Indeed, if verifiable weapons had been found instead, the clamour in the scientific journals would have been enormous. That this clamour never happened should be telling you something important.
Plus, your attempt to peddle the tired old creationist trope, that absence of evidence purportedly doesn't mean evidence of absence, apart from being tiresome, falls flat on its face courtesy of those tool discoveries with respect to Australopithecus. Scientists didn't think that Australopithecus utilised tools of any sort until 2010, when those tools were discovered. Once those tools were discovered, the scientists changed their view in accordance with the new data. This is how real science is done, not by making up fantasy scenarios and then pretending that these constitute established fact, despite a wealth of DATA saying otherwise.
Here's a clue for you, JayJay. We're not going to accept your blind, evidence-free assertions as fact, just because you keep robotically parroting them during your creationist Energiser Bunny act. We're going to wait for DATA to change our minds, not your apologetic duplicity and discoursive intransigence.
Oh look, here we go again, with the duplicitous hand-waving away of an evidentially supported phenomenon, by summarily dismissing it as a "trope", whilst ignoring totally that your fantasy assertions have ZERO evidential support. Sexual selection has been documented to be at work in hundreds, if not thousands of species, and it's manifestly at work in humans too, as any time spent in a nightclub watching testosterone-fuelled males trying to get laid by picky females will inform anyone not wearing your ideological blinkers.
Except that oops, long canines in primates are frequently observed to be the PRODUCTS of the very same sexual selection you duplicitously tried to hand-wave away above.
Except that, oh wait, I destroyed your quote mining of the Cheney et al paper back on page 29 of this thread in this post, and repeated said demolition in this subsequent post, where I reminded everyone of what the paper actually stated.
In your dreams ...
Different social organsiation. As Lovejoy explained in that 2009 paper I've just presented. Next?
No. A different social organisation, involving much lower levels of male-male combative intensity, made them superfluous to requirements.
Bollocks. Once again, NO RELIABLY DETERMINABLE WEAPONS HAVE BEEN FOUND DATING BACK MORE THAN 100,000 YEARS. Game. Fucking. Over.
Bollocks. Once again, NO RELIABLY DETERMINABLE WEAPONS HAVE BEEN FOUND DATING BACK MORE THAN 100,000 YEARS. Game. Fucking. Over.
Drivel. Every nightclub on the planet contains evidence telling us that you're posting drivel.
No it isn't an "inference", it's a fucking fantasy. Oh wait, once again, NO RELIABLY DETERMINABLE WEAPONS HAVE BEEN FOUND DATING BACK MORE THAN 100,000 YEARS. Game. Fucking. Over.
Because that's what you're peddling here, a fucking fantasy. When is this going to sink in?
Three words. Lithic wear analysis. Which I'm going to keep humiliating you with. You know, the [b]rigorous method of determining what use a tool was put to? And which you've avoided the way right wing politicians avoid telling the truth?
OK, we now say goodbye to Part 1. Part 2 follows shortly.
As this is a long post, it's a two parter. Welcome to Part 1.
Well given that much of your output has repeatedly been demonstrated to be lies and bullshit, courtesy of the DATA telling us this, I'm merely taking this fact on board in advance before going on to demolish more of the lies and bullshit you post. Much of which turned out to be lies and bullshit once again once I addressed it. In short, JayJay, if you don't want your output to be regarded as duplicitous and worthy only of scorn and derision as a result, then there's a simple answer - instead of engaging in synthetic whingeing and bleating about post style when you're caught in flagrante delicto, stop posting manifest falsehoods and trying to pass them off as fact. Or is this simple answer too difficult for you even to acknowledge the existence of, let alone try to implement?
Oh, and as for a "reasonable" discussion, JayJay, my idea of a "reasonable" discussion, is one in which proper evidence is presented to support ideas, not made up shit. By that criterion, your entire posting career here has been wholly unreasonable, yet, instead of merely whingeing and bleating about this, I and the others here have exercised the diligent effort to demonstrate that your posts are lies and bullshit from start to finish. This is how discourse is conducted by grown ups.
Jayjay4547 wrote:Calilasseia wrote:Jayjay4547 wrote:But my point is(a) that the lack of fangs in Australopithecus should have immediately created the inference that they instead used hand held weapons that made them dangerous to attack.
Bollocks. First of all, there are plenty of organisms that don't possess big canines, and which don't use tools.
Armadillos would be an example of animals that don’t possess big canines and don’t use tools.
Oh I can think of plenty of others. How about this for a nice selection?
Starting with the placental mammals, we'll run with, say, Heterohyrax brucei, shall we?
Or how about Rhyncocyon petersi, the Black & Rufous Elephant Shrew?
Or Galeopterus variegatus, the Colugo or Sunda Flying Lemur?
Or how about the Dugong, Dugong dugon?
Or for that matter, its close relative, the Antillean Manatee, Trichechus manatus?
Oh, but we forgot the marsupials! Have a Koala Phascolarctos cinereus, for example:
Or the Sugar Glider, Petaurus breviceps:
Or Antechinus stuarti, one of several marsupial mice in the Genus, famous for its males literally shagging themselves to death when reaching sexual maturity:
Or the unique Dromiciops gliroides, the sole living representative of the Microbiotheria (which also has no less than 50 teeth):
Then we have the Platypus, Onithorhynchus anatinus, a unique still-extant Monotreme :
What about other Vertebrates? Try this one - a female Black-Chinned Hummingbird, Archilochus alexandri ... there are 338 species of hummingbird known to science currently:
Or the Red-Crested Turaco, Tauraco erythrolophus:
Or the Bar-Throated Minla, Minla stigula:
Or perhaps, moving on to reptiles, we could choose this one, Cercosaura argulus :
Here's another nice organism lacking canines or tool usage. Say hello to Forcipiger longirostris from Hawaii:
Or how about this one? Say hello to Corydoras panda, which apart from being a species I've bred in the aquarium, also lacks oral dentition of any sort:
Perhaps a better candidate would be Beaufortia kweichowensis, one of the hillstream loaches, which again lacks oral dentition of any sort, and for that matter lacks any form of armour, being a scaleless fish? Here it is:
Just for the record, the Cobitidae, Balitoridae, Nemacheilidae and Gastromyzontidae, are all scaleless fishes. As are many of the catfishes in the Order Siluriformes. In the case of the loaches, the taxonomy is somewhat in flux at the moment, while Kottelat performs his major revision of the entire clade. But oops, we left out the INvertebrates! Let's remedy this shall we?
Well start with Carausius morosus, the Indian Stick Insect:
Or how about Cethosia nietneri?
(I won't bother with any more insects, as I have something like 40,000 photos of these on my hard drive as it is ...
Here's another invertebrate, in the form of Anadenobolus monilicornis :
Then there's this one, Daphnia pulex :
Or perhaps Argulus foliaceus is more to your liking?
Or how about Chromodoris annae, one of many species of nudibranch?
Or Elysia crispata, if you prefer something a little more rococo in appearance?
Or Pteraeolidia ianthina?
Or try Schmidtea mediterranea, which is being added to the list of laboratory model organisms by genetics researchers ...
Or how about Caenorhabditis elegans, which has been so informative to scientists in recent decades?
Finally, how about these sponges?
I was going to toss into the mix this beast, namely one of the Warnowiids:
but this takes us outside the Metazoa, because dinoflagellates are algae, and therefore plants, though in the case of Warnowiids, they're not only mobile but have eyes. Just to illustrate how diverse the biosphere really is, and that viewing its contents in the same manner as a three year old's pop-up book on Noah's Ark will merely elicit much pointing and laughing. Which provides much of the reason for me carpet bombing you with all those photos, because all too often, your view of the biosphere is that of a three-year-old's pop up book on Noah's Ark. Though perhaps not as floridly so as American creationists, whose thinking about the biosphere consists of thinking that the rest of the species on the planet were put here to provide them with burgers and fries. But a timely reminder that we're not so fucking special after all is apposite here.
Moving on ...
Jayjay4547 wrote:The issue isn’t about “tools” in general, but specifically about hand held weapons, used to make a species dangerous to attack.
Except that once again, NO EVIDENCE OF SUCH WEAPONS OLDER THAN 100,000 YEARS BEFORE PRESENT HAS EVER BEEN FOUND. Game. Fucking. Over.
Jayjay4547 wrote:For that, an animal would need hands capable of grasping a weapon, means to carry a weapon around, it would need to be of a certain absolute size where kinetic weaponry can be effective, and a certain size relative to predator.
It would also need to be capable of formulating the idea. Except that oops, we have NO EVIDENCE that hominids of any taxonomic designation, formulated the idea of developing hand held weapons until just 100,000 years ago. Game. Fucking. Over.
Jayjay4547 wrote:Australopithecus satisfied those criteria
Except that, oops, NO EVIDENCE OF HAND HELD WEAPONS FASHIONED BY AUSTRALOPITHECINES EXISTS. Game. Fucking. Over.
Jayjay4547 wrote:and higher primates generally are near the edge of it, shown by Kortlandt’s 1960s experiments with chimps and a dummy leopard
Which I've already questioned ruthlessly when you first posted this. Funny how you never bothered to answer this with substance.
Jayjay4547 wrote:and by Boesch’s account of chimps taunting a cornered leopard using sticks.
Oh wait, what do all of these instances have in common? Social organisation. Which the DATA tells us appears to be required to be in place for a protracted period of time first.
Jayjay4547 wrote:We have been over those a few times.
And I demolished your made up shit then as well.
Jayjay4547 wrote:Anyway I came across a cute supporting anecdote the other day, about someone who, hoping to protect his car bonnet and windscreen wipers from damage by baboons in the Cape Point nature reserve, left a rubber snake on the dashboard. When he got back to his car he found the windscreen had been smashed by baboons, using stones.
Know this for a fact, does he? Was this actually recorded happening?
Oh, you mean you're going to make more shit up on this subject, despite me having presented the original scientific paper describing the species, in which the canines were determined to be no bigger than 11 mm? Oh that's going to be fucking hilarious when you do.
Jayjay4547 wrote:but really its no more needed to find tools tied to an ancestor of Australopithecs, than one needs to find bones stuck in the teeth of T.rex to figure out it ate meat.
Except that, oh wait, scientists didn't just fucking ASSUME that T.rex teeth were indicative of a carnivorous trophic niche, they set about acquiring DATA to establish this. Starting with a hell of a lot of time spent making comparative anatomy a rigorous discipline. Then, lo and behold, they also found fossils of prey organisms bearing bone marks consistent with T. rex attack.
Jayjay4547 wrote:Palaeontologists had not trouble figuring that out, long before they found marks left by those animals.
Oh wait, what were those two words I posted above again? Comparative anatomy. Which had been in place for some 100 years or so before palaeontology really took off. Which in turn became possible thanks to Linnaeus launching the taxonomic enterprise, and encouraging concentration on detail.
Jayjay4547 wrote: On top of that, Your “3 million year history” is more correctly a 3 million year GAP between the date of one fossil actual or near ancestor, and Australopithecus.
Er, wrong. Apart from the fact that gaps are a known creationist fetish, we've plenty of evidence for species persisting for far longer periods than 3 million years, and indeed, 3 million years would be an entirely reasonable time scale for a transition from Sahelanthropus to Australopithecus. You're also forgetting Ardipithecus, another pre-Australopithecine hominid Genus, with A. ramidus dating to 4.4 million years before present, and A. kadabba, dating to 5.6 million years before present. None of these organisms exhibited any propensity for tool use either.
It's time for another scientific paper, namely:
Re-Examining Human Origins In Light Of Ardipithecus ramidus by C. Owen Lovejoy, Science, 326: 74-74e8 (2nd October 2009) [Full paper downloadable from here]
Lovejoy, 2009 wrote:Referential models based on extant African apes have dominated reconstructions of early human evolution since Darwin’s time. These models visualize fundamental human behaviors as intensifications of behaviors observed in living chimpanzees and/or gorillas (for instance, upright feeding, male dominance displays, tool use, culture, hunting, and warfare). Ardipithecus essentially falsifies such models, because extant apes are highly derived relative to our last common ancestors. Moreover, uniquely derived hominid characters, especially those of locomotion and canine reduction, appear to have emerged shortly after the hominid/chimpanzee divergence. Hence, Ardipithecus provides a new window through which to view our clade’s earliest evolution and its ecological context. Early hominids and extant apes are remarkably divergent in many cardinal characters. We can no longer rely on homologies with African apes for accounts of our origins and must turn instead to general evolutionary theory. A proposed adaptive suite for the emergence of Ardipithecus from the last common ancestor that we shared with chimpanzees accounts for these principal ape/human differences, as well as the marked demographic success and cognitive efflorescence of later Plio-Pleistocene hominids.
From that paper in more detail:
Lovejoy, 2009 wrote:Ardipithecus ramidus. Ardipithecus ramidus now reveals that the early hominid evolutionary trajectory differed profoundly from those of our ape relatives from our clade’s very beginning. Ar. ramidus was already well-adapted to bipedality, even though it retained arboreal capabilities (19–25). Its postcranial anatomy reveals that locomotion in the chimpanzee/human last common ancestor (hereafter the CLCA) must have retained generalized above-branch quadrupedality, never relying sufficiently on suspension, vertical climbing, or knuckle walking to have elicited any musculoskeletal adaptations to these behaviors (26–28).
Moreover, Ardipithecus was neither a ripe-fruit specialist like Pan, nor a folivorous browser like Gorilla, but rather a more generalized omnivore (19, 25). It had already abandoned entirely the otherwise universal sectorial canine complex (SCC), in which the larger, projecting upper canine is constantly honed by occlusion against the lower third molar of anthropoid primates (25), demonstrating that the large, projecting, interlocking, and honing male canines of apes had been eliminated before the dawn of the Pliocene and before the emergence of the dentognathic peculiarities of Australopithecus. What’s more, it appears to have been only slightly dimorphic in body size (25). Finally, the environmental context of Ardipithecus suggests that its primary habitat was not savanna or grassland, but instead woodlands (26–28).
In retrospect, clues to this vast divide between the evolutionary trajectories of African apes and hominids have always been present. Apes are largely inept at walking upright. They exhibit reproductive behavior and anatomy profoundly unlike those of humans. African ape males have retained (or evolved, see below) a massive SCC and exhibit little or no direct investment in their offspring (their reproductive strategies rely primarily on varying forms of male-to-male agonism). Although they excel at some cognitive tasks, they perform at levels qualitatively similar to those of some extraordinary birds (29, 30) and mammals (31). The great apes are an isolated, uniquely specialized relict species surviving today only by their occupation of forest refugia (32). Even their gut structure differs substantially from that of humans (33).
The author provides this in the preamble article preceding the paper:
Lovejoy, 2009 wrote:Loss of the projecting canine raises other vexing questions because this tooth is so fundamental to reproductive success in higher primates. What could cause males to forfeit their ability to aggressively compete with other males? What changes paved the way for the later emergence of the energy thirsty brain of Homo? Such questions can no longer be addressed by simply comparing humans to extant apes, because no ape exhibits an even remotely similar evolutionary trajectory to that revealed by Ardipithecus.
When the likely adaptations of early hominids are viewed generally rather than with specific reference to living chimpanzees, answers to such questions arise naturally. Many odd hominid characteristics become transformed from peculiar to commonplace. Combining our knowledge of mammalian reproductive physiology and the hominid fossil record suggests that a major shift in life-history strategy transformed the social structure of early hominids. That shift probably reduced male-to-male conflict and combined three previously unseen behaviors associated with their ability to exploit both trees and the land surface: (i) regular food-carrying, (ii) pair-bonding, and (iii) reproductive crypsis (in which females did not advertise ovulation, unlike the case in chimpanzees). Together, these behaviors would have substantially intensified male parental investment—a breakthrough adaptation with anatomical, behavioral, and physiological consequences for early hominids and for all of their descendants, including ourselves.
Moving back to the paper itself, we have this:
Lovejoy, 2009 wrote:Adaptive suites. An alternative to referential modeling is the adaptive suite, an extrapolation from optimization theory (35). Adaptive suites are semiformal, largely inductive algorithms that causally interrelate fundamental characters that may have contributed to an organism’s total adaptive pattern. One for the horned lizard (Phyrnosoma platyrhinos) of the southwesten U.S. serves as an excellent example (Fig. 1) (36, 37). For this species, the interrelation between a dietary concentration on ants and its impact on body form imply, at first counterintuitively, that elevation of clutch size and intensification of “r” strategy (maximize the number of offspring by minimizing paternal care) are the ultimate consequences of this specialization (35–37).
Such character and behavioral interdependencies can have profound consequences on evolutionary trajectory, as demonstrated by the equally notable differences in clutch size in the common leopard frog (3500 to 6500 eggs) versus those of numerous species of so-called poison dart frogs [typically less than 30 eggs; Table 1 (38)]. To enhance survival of their (as yet) nontoxic offspring, the latter engage in relatively intense male parental investment, a shift that has had a profound adaptive impact on their entire life-history strategy.
The effective power of adaptive suites is demonstrable by their explanatory success. A virtually identical character constellation to that of the horned lizard has been discovered in an unrelated Australian ecological vicar, Moloch horridus (37), which is also an ant specialist. Even given such unexpected consilience, however, adaptive suites are obviously speculative, even for living organisms. In addition, for extant species, the processes by which current characters have emerged are also necessarily hidden in the past and, therefore, are no more accessible than for extinct taxa. Nevertheless, adaptive suites can serve as organizational procedures by which to examine evolutionary processes with increasing acumen. Of further benefit is the fact that they often pose novel testable hypotheses that might not have arisen without them.
Many key human specializations are related to our reproductive physiology and anatomy; human reproduction is as extraordinary as our dentition, locomotion, and encephalization (39). Although it remains possible that such uniqueness emerged only during the Pleistocene, this is less likely in light of Ardipithecus, which shows very early evidence of a major social transformation (25). Moreover, it is the modern African apes that are most derived in many characters, whereas those which are specialized in human evolution (SCC elimination, bipedality) are now known to have been present near the origin of our clade. Our massive brains are obviously a Pleistocene development, but they are also probably sequelae to other major shifts now more fully recorded in the earlier fossil record. It is therefore possible, even likely, that many physiologies and soft tissue features that do not fossilize may have also evolved early in hominid evolution. If so, why were these characters exaptive to our advanced cognition and singular demographic success?
Notwithstanding the revelations now provided by Ardipithecus, it should be noted that extensive studies of African apes and other primates have provided likely details of the sociobehavioral context from which hominids most likely first emerged (1, 11). These details were presumably present in the last common ancestor we shared with the African apes, and they almost certainly included aspects of great ape demography and social behavior, including male philopatry (males remain in their natal group), female exogamy (females transfer from natal group at sexual maturity), and prolonged inter-birth intervals, all cardinal characters of an intense “k” (maximized parental care of few offspring) reproductive strategy (32, 40). Moreover, investigations of the behavior of other living primates now provide a wealth of information that allows many contextual details of earliest human evolution to be reasonably hypothesized.
Sperm competition. Two key factors dominate anthropoid reproductive behavior and are therefore diagnostic of socio-sexual structure: (i) sperm competition and (ii) male-to-male competition for mates. Various anatomical correlates distinguish monogamous or single male primates from other species whose males engage in sperm competition. Among the most obvious is the much higher ratio of testes volume to body mass. Human ratios are generally similar to those of monogamous gibbons and solitary orangutans, but the ratios are three times higher in Pan (41, 42) and other sperm-competing species such as Brachyteles (43). Moreover, human testes are most similar to those of gibbons with respect to their higher proportion of intertubular (nonseminiferous) tissue (42). Mammalian sperm competition is generally accompanied by elevated ejaculate quality (44), which is also notably poor in humans. In Homo sapiens, the absolute rate of sperm production is only about 20% that of much smaller rhesus macaques (45). Another measure, spermatogenesis efficiency (daily sperm production per gram of testes), “varies from about 2.65 × 107 in rabbits to <0.6 × 106 in humans” (46). [NOTE: figure corrected in accordance with erratum at the end of the paper]. The estimated corresponding value in chimpanzees is greater than that of humans by two orders of magnitude (42).
The muscular coat of mammalian vasa deferentia can reasonably be regarded as a correlate of sperm transport rate during sexual stimulation. It is substantially thicker in chimpanzees than in humans or orangutans (47). The seminal vesicles of some monogamous primates are inconspicuous, whereas those of multimale (i.e., ovulating females usually mate with multiple males) macaques and chimpanzees are large; those in humans are only of moderate size (39). Whatever the social caveats may be, human ejaculatory rates (along with those of the monogamous genera Aotus and Symphalangus[i]) are lower than those of 20 primate species (including [i]Pan and Gorilla) by one order of magnitude (48, 49), and human sperm counts decrease at ejaculation frequencies of >4 per week (50).
Human sperm midpiece volume, which reflects mitochondrial density and motility, falls in the lowest quartile of 21 primate species examined (51). Especially important is the coagulating reaction between some seminal proteins and prostate vesiculase (52). This coagulum, which blocks penetration of competing sperm by forming a vaginal plug, characterizes primates that robustly spermcompete (e.g., Ateles, Brachyteles, Macaca, Pan). This reaction is absent in humans and common marmosets, whose ejaculates are merely gelatinous (53).
The structures of semenogelins I and II (SEMGI and SEMGII) (primary plug coagulates) illuminate the natural history of vaginal plugging. SEMGI suggests a selective sweep in chimpanzees and conversion to a pseudogene in gorillas; humans exhibit neither (52). Together, these data strongly suggest that the social structure in earlier hominids is unlikely to have been typically multimale. This conclusion is supported by recent analyses of primate immune systems, which compared basal white blood cell counts among primates with respect to the likely number of sexual partners as determined by social system (female mating promiscuity). Results showed that “humans align most closely with the [single male] gorilla ... and secondarily with … [the] monogamous gibbon” [(54), p. 1170].
Humans have the least complex penis morphology of any primate. Complexity is generally associated with multimale social structure (47), and humans lack keratinous penile surface mechanoreceptors that may promote rapid ejaculation that is common in many primates. Finally, humans are the only catarrhine without an os baculum (39).
Competition for mates. If they did not spermcompete, did early hominid males instead compete for single or near-solitary control of female groups? The cardinal indicator of male-to-male agonism in hominoid primates is the SCC. It is regularly employed during both territory defense and dominance disputes. Hominids are often characterized as having reduced canine dimorphism (55). Such reduction is only a secondary consequence of the primary hominid character, which is elimination of the SCC in its entirety. The SCC is not male-limited; that is, it is always expressed in both sexes of all anthropoids, even in species with reduced dimorphism (e.g., some New World atelines). Although females may express the SCC for advantage in conflicts with other females, they principally express its underlying structure themselves because amplification in their male offspring (presumably by androgens or reduced estrogens) enhances their fitness. Hylobatid canine monomorphism is sometimes erroneously confused with that of hominids, but gibbons evolved amplification of the female canine. Ar. ramidus shows that elimination of the SCC in hominids is unique among all higher primates and occurred long before Australopithecus.
A frequent explanation of canine reduction (and bipedality) is that hand-held weapons replaced the SCC (56, 57). But if male-to-male agonism had been fundamental to early hominid fitness, what selective agency would have reduced its signature character? Additional human attributes belie the improbability of the weapons argument. An absence of sperm competition in gorillas and orangutans is accompanied by a dramatically reduced testes size and the elimination of a free scrotum (their testes are more judiciously sequestered in a post-penial bulge) (42, 58). In contrast, not only are human scrota more pendulous than even those of chimpanzees (58), but bipedality makes them extraordinarily vulnerable during upright combat (59). It seems illogical to attribute habitual uprightness to weapons that would demand even greater selection for testes sequestration than is present in other primates [which target them with their functional SCCs (60)].
Available evidence now suggests that the loss of the SCC was, as is theoretically most likely, a social adaptation. This evidence, derived from Ardipithecus, includes the following (25): (i) Change in the more socially important upper canine preceded that in the lower, (ii) progressive shape modification made the canine not only smaller but less weaponlike in form, (iii) male canines erupted relatively earlier than in large-canined species with high male-to-male agonism, making this event less likely to have represented a social signal of male maturity, and (iv) all of these changes took place within a dietary context that preceded any of the profound changes seen in later hominid dentitions.
Humans are also unique among primates in lacking vocal sacs, which play a major role in the territoriality of all apes. Though there are no current means by which to judge the evolutionary history of the hominoid vocal apparatus (61), it does have potential developmental interactions with basicranial patterning, including an impact on location of the foramen magnum. The dramatic anterior translation of this foramen during the Plio-Pleistocene is almost certainly a corollary of cerebral reorganization and/or expansion (62). However, early hominid vocal apparatus reduction may have influenced initial differential trajectories of cranial form, currently only just detectable in P. paniscus and Ar. ramidus (29). Both cerebral reorganization and facial patterning are clearly central elements of that trajectory, and early reduction of vocal tract mass is thus a potential modulating factor, particularly because it is a possible social corollary of loss of the SCC.
It has long been argued that Australopithecus was unusually dimorphic in body size, implying a largely single-male group structure, but this hypothesis has been biased by comparisons of temporally and geographically disparate samples (63). Of greater importance are (i) the absence of any useful correlation between body-size dimorphism and social structure in hominoids, because both chimpanzees and gorillas exhibit intense male-to-male agonismbut exhibit opposite polarities in skeletal dimorphism (63); and (ii) the fact that male body size in many primates is not associated with competition for mates. Rather, it is equally likely to be an ecological specialization derived from reduced size of females (64) and/or male enlargement by selective agencies unrelated to mate acquisition. In any case, Ar. ramidus now transcends the debates over dimorphism in early Australopithecus because available samples indicate that it was minimally dimorphic, suggesting that this was the primitive hominid condition (19, 25) and that dimorphism increased in later hominids (see below).
From the diagram on page 74e4, we have this:
Lovejoy, 2009 wrote:(Right column) The demographic dilemma (32, 79) generated by intensified K selection. A solution for a hominoid confronting such selective forces is elaboration of sex-for-food exchanges observed in chimpanzees and bonobos. These and other elements shared with Pan acted as possible “social catalysts” [highlighted in red (e.g., copulatory feeding, extractive foraging, male-male patrols)]. Increased male body size and enhanced male-to-male cooperation in Au. afarensis reduced mortality during distance foraging by multiple-male patrols (whose role was optimal foraging rather than territory defense). This culminated in savanna scavenging, primitive lithics for meat acquisition, marrow extraction, and cooperative hunting in Homo. This profound economic shift selected for advanced adaptations to bipedality, further enhanced social cohesion (reduced same-sex agonism in both sexes), increased energy available for parenting (and alloparenting), promoted survivorship and reduced birth spacing, and elevated the selective ceiling acting against metabolically expensive tissues (e.g., the brain).
The paper continues as follows:
Lovejoy, 2009 wrote:Reproductive crypsis: the most unique human character. Elimination of the SCC and the ecological context of Ardipithecus at Aramis, Ethiopia, and earlier sites are consistent with the inference that male provisioning via resource transport (and concomitant terrestrial bipedality) antedated 4.4 Ma. Might such behaviors have first evolved nearer the base of the hominid clade? An obvious issue with the hypothesis outlined above is that Pan males prefer females with prominent signs of active ovulation (estrus). If minimal ovulatory signaling in the earliest hominids was primitive (as it is presumably in Gorilla), why did hominid females not prolong and intensify such signs so as to encourage sex-for-food exchanges?
First, the extreme ovulatory-related displays in Pan appear to be derived, because they are associated with comparatively unique molecular signatures of accompanying adaptations (such as proteins necessary for vaginal plugging) absent in other hominoids, as well as appropriately specialized penile morphology. Second, it is unlikely that copulation offered by a female would be rejected by a male—this would be counterproductive given the substantial variability of the primate menstrual cycle and the rarity of hominoid females available for impregnation. Furthermore, habitual provisioning of a target female, even while still lactating for a dependent infant, would still make the repeatedly attendant male most likely to sire any successive offspring upon first reinstatement of ovulatory cycling.
The latter point is critical. One of the most frequently cited objections to male provisioning in early hominids is the problem of cuckoldry during times that males would have been separated from a selected mate while in search of food (83, 84). But ovulation in hominoids is an exceptionally rare event, and it probably occurs only after extensive, 3- to 4-year-long periods during which female lactation amenorrhea prevents it. Male provisioning of rarely but obviously fertile females would enhance his fitness by several means: (i) Regular copulation would probabalistically establish an attendant male as the most likely to sire the target female’s succeeding offspring, provided that his mate did not “advertise” her ovulation and/or solicit multiple copulations. (ii) Repeated provisioning would accelerate reinstatement of ovulation by replenishing fat stores depleted by lactation. (iii) Accidental or pathological death of her dependent offspring (a not infrequent event) would also reinitialize ovulation, and selection would obviously favor habituation with nonaggressive males not predisposed to infanticide, which is already unlikely because of philopatry.
To prevent cuckoldry, male provisioning within the context of a multimale group therefore requires selection of females with reproductive crypsis. That is, males could only succeed by provisioning mates with self-crypsis; they would otherwise be unprotected from female copulation with more dominant/aggressive males while ovulating. Broadly (but not entirely) nonovulatory copulation, as in Brachyteles (69, 70), would permit prolonged exclusivity in operational pair bonds, especially when provisioning males showed preference for females who were not observed to copulate with other males (85). In this context, it is therefore relevant that human females do not externally advertise ovulation [other hominoids exhibit some degree of ovulatory swelling, even if minimal (86)] and also fail to exhibit its substantial physiological self-perception, despite moderately elevated proceptivity during ovulation (39, 87).
So the author presents the dynamics of reproduction within a social group as being primary determinants of the requisite features, not your fantasy acquisition of weapons, and presents data supporting this linkage.
From the conclusion:
Lovejoy, 2009 wrote:Conclusion. As Au. afarensis was progressively revealed during the 1970s, its anatomy and antiquity still permitted a possible chimpanzee-like CLCA. Many models of human origins, largely referential, employed this perspective. Previous nonreferential attempts (32, 79) argued that only major changes in the social behavior of Au. afarensis and its ancestors could satisfactorily account for its unique combination of postcranial anatomy and unusual demographic success. The tempo and mode of such hypothetical earlier evolutionary events, however, have remained shrouded from our view. This has led to rejection of the hypothesis by many who preferred the comparative comfort and safety of more referential accounts.
Even as its fossil record proliferated, however, Australopithecus continued to provide only an incomplete understanding of hominid origins. Paradoxically, in light of Ardipithecus, we can now see that Australopithecus was too derived— its locomotion too sophisticated, and its invasion of new habitats too advanced—not to almost entirely obscure earlier hominid evolutionary dynamics.
Now, in light of Ar. ramidus, there are no longer any a priori reasons to suppose that acquisition of our unique reproductive anatomy and behavior are unconnected with other human specializations. The evidence is now conclusive: Elimination of the SCC occurred long before the eventual dentognathic hypertrophy of Australopithecus, and long before the likely horizon at which sufficient reliance on tool use would have encouraged abandonment of food and/or safety in the arboreal substrate. It is far more likely that our unique reproductive behavior and anatomy emerged in concert with habituation to bipedality and elimination of the SCC (Fig. 2). It is also now equally clear that Pan’s specialized reproductive constellation has been driven by an entirely different locomotor and dietary history.
We currently know very little about the postcranium of hominids older than Ar. ramidus (e.g., Sahelanthropus, Orrorin) (105, 106). More fossils will further advance our understanding of the CLCA, and we anxiously await their discovery. Meanwhile, the opportunity of devising adaptive suites for both species of Pan and for Gorilla—grounded in hypotheses generated in light now thrown on the gorilla/chimpanzee/human last common ancestor and CLCA by Ar. ramidus as to their locomotion, diet, and social behavior—is an intriguing prospect whose alternative outcomes will probably provide additional revelations.
When viewed holistically, as any adaptive suite requires, the early hominid characters that were probably interwoven by selection to eventually generate cognition now seem every bit as biologically ordinary as those that have also affected the evolution of lizards, frogs, voles, monkeys, and chimpanzees. Comparing ourselves to our closest kin, it is somewhat sobering that the hominid path led to cognition, whereas that leading to Pan, our closest living relatives, did not, despite the near-synonymy of our genomes.
As Darwin argued, the ultimate source of any explication of human acumen must be natural selection (78). The adaptive suite proposed here provides at least one evolutionary map by which cognition could have emerged without reliance on any special mammalian trait. The perspective offered by Ardipithecus suggests that our special cognitive abilities derive from a unique earlier interplay of otherwise commonplace elements of locomotion, reproductive biology, neurophysiology, and social behavior. In retrospect, we are as ordinary as corvids (107) and voles (108), although we are much more fortunate, if self-cognition is deemed fortunate. We should never have doubted Darwin in his appreciation that the ultimate source of our matchlessness among mammals would prove commonplace when knowledge became sufficiently advanced. Ar. ramidus now enhances that knowledge. Even our species-defining cooperative mutualism can now be seen to extend well beyond the deepest Pliocene.
Looks like once again, your assertions have received a thorough fisking from real science.
Jayjay4547 wrote:You demand for evidence, a tool found in that gap, to be positively tied to Sahelanthropus, for lack of which you pronounce my “fantasy” destroyed by “DATA”.
Oh wait, what happened when tools were found that could be reliably associated with Australopithecus? Analysis of those tools revealed them to be food preparation tools. For a long time, even these were unknown to be associated with this taxon. But given the ubiquity of feeding related tools throughout the Eumetazoa, it's not surprising to those of us who paid attention in biology class, that the first tools to be found in association with Australopithecus should turn out to be tools of this sort. Indeed, if verifiable weapons had been found instead, the clamour in the scientific journals would have been enormous. That this clamour never happened should be telling you something important.
Plus, your attempt to peddle the tired old creationist trope, that absence of evidence purportedly doesn't mean evidence of absence, apart from being tiresome, falls flat on its face courtesy of those tool discoveries with respect to Australopithecus. Scientists didn't think that Australopithecus utilised tools of any sort until 2010, when those tools were discovered. Once those tools were discovered, the scientists changed their view in accordance with the new data. This is how real science is done, not by making up fantasy scenarios and then pretending that these constitute established fact, despite a wealth of DATA saying otherwise.
Here's a clue for you, JayJay. We're not going to accept your blind, evidence-free assertions as fact, just because you keep robotically parroting them during your creationist Energiser Bunny act. We're going to wait for DATA to change our minds, not your apologetic duplicity and discoursive intransigence.
Jayjay4547 wrote:Calilasseia wrote:Jayjay4547 wrote: It’s nonetheless true that the long sharp canines of other higher primates do make them dangerous attack. They are ferocious biters.
Oh wait, how often have you been presented with DATA, JayJay, demonstrating that a significant proportion of the use of those canines, focuses on competition between rivals of the same species for a highly desirable resource?
Indeed, how many primate species exhibit manifest sexual dimorphism in this regard, which makes NO sense whatsoever in the light of your obsessive concentration on defensive biting as purportedly the sole reason for this dentition, even before we factor in such things as diet? Because one of the elementary thoughts that immediately crosses the mind of anyone who has paid attention in biology class, is that a defensive capability of this sort would be useful to members of a given species regardless of gender, if a pressing need accompanied by a measurable selection pressure was present for said capability. The mere fact that something as functional as dentition, undergoes gender biased modification, tells those of us who paid attention in biology class, that your obsessive fantasising about defensive biting is precisely that, because modification of this sort would not affect significantly, a feature already subject to an alternative strong selection pressure.
To repeat, I’ve sidestepped that whole trope about sexual selection
Oh look, here we go again, with the duplicitous hand-waving away of an evidentially supported phenomenon, by summarily dismissing it as a "trope", whilst ignoring totally that your fantasy assertions have ZERO evidential support. Sexual selection has been documented to be at work in hundreds, if not thousands of species, and it's manifestly at work in humans too, as any time spent in a nightclub watching testosterone-fuelled males trying to get laid by picky females will inform anyone not wearing your ideological blinkers.
Jayjay4547 wrote:by just sticking to the point that the long sharp canines of other higher primates make them dangerous to attack, which is significant for their ecology.
Except that oops, long canines in primates are frequently observed to be the PRODUCTS of the very same sexual selection you duplicitously tried to hand-wave away above.
Jayjay4547 wrote:Consider that video clip I showed about humans very circumspectly trying to attack a group of baboons occupying and then rifling a car. The large male on top of the car intimidated the men, keeping them away until the baboons had got what they wanted. The observation by Cheney and others that all the members of troop get involved in a mobbing attack doesn’t contradict that the sexually dimorphous male primates are the ones who can damage- and everyone knows that instinctively.
Except that, oh wait, I destroyed your quote mining of the Cheney et al paper back on page 29 of this thread in this post, and repeated said demolition in this subsequent post, where I reminded everyone of what the paper actually stated.
Jayjay4547 wrote:Here is a problem for you.
In your dreams ...
Jayjay4547 wrote:Suppose that primate sexual dimorphism in canines is driven by sexual selection. Why is it then that men and ancestral Australopithecus males lack long canines? Why is it that we men have inherited such girly skulls?
Different social organsiation. As Lovejoy explained in that 2009 paper I've just presented. Next?
Jayjay4547 wrote:Were Australopithecus utterly above such crude competition for the possession of females, as Darwin put it?
No. A different social organisation, involving much lower levels of male-male combative intensity, made them superfluous to requirements.
Jayjay4547 wrote: Seems to me that the Australopithecus males competed using stick fights just like young tribal men today.
Bollocks. Once again, NO RELIABLY DETERMINABLE WEAPONS HAVE BEEN FOUND DATING BACK MORE THAN 100,000 YEARS. Game. Fucking. Over.
Jayjay4547 wrote:And those skills were turned against predators, making a hominin troop dangerous to attack.
Bollocks. Once again, NO RELIABLY DETERMINABLE WEAPONS HAVE BEEN FOUND DATING BACK MORE THAN 100,000 YEARS. Game. Fucking. Over.
Jayjay4547 wrote:Intraspecific competition in primates and access to females is just a cruel trick by “nature” to keep testosterone filled individuals from breeding out of the gene pool.
Drivel. Every nightclub on the planet contains evidence telling us that you're posting drivel.
Jayjay4547 wrote:Calilasseia wrote: But this is another piece of DATA you keep ignoring whilst trying to peddle your sad little fantasy as purportedly constituting fact,
It’s not a fantasy, not sad, or even little. It’s a strong INFERENCE that Australopithecus used hand held weapons to make them dangerous to attack.
No it isn't an "inference", it's a fucking fantasy. Oh wait, once again, NO RELIABLY DETERMINABLE WEAPONS HAVE BEEN FOUND DATING BACK MORE THAN 100,000 YEARS. Game. Fucking. Over.
Jayjay4547 wrote:You used that word “fantasy” 16 times in your post.
Because that's what you're peddling here, a fucking fantasy. When is this going to sink in?
Jayjay4547 wrote:Calilasseia wrote: regardless of how much DATA destroys that fantasy. DATA such as:
1: NO hominid weapons older than 100,000 years have ever been found;
When I look at your so called DATA it turns out to date the appearance of hafted weapons used for hunting by fully modern Homo sapiens. That’s only a bit less ridiculous than demanding to see a rifle before admitting weaponry.
Three words. Lithic wear analysis. Which I'm going to keep humiliating you with. You know, the [b]rigorous method of determining what use a tool was put to? And which you've avoided the way right wing politicians avoid telling the truth?
OK, we now say goodbye to Part 1. Part 2 follows shortly.
Signature temporarily on hold until I can find a reliable image host ...
-

Calilasseia - RS Donator
- Posts: 22642
- Age: 62
- Country: England
Re: How atheist ideology messed up the human origin story
#1010 ![]() by Calilasseia » Dec 15, 2015 2:46 pm
by Calilasseia » Dec 15, 2015 2:46 pm
Welcome to Part 2.
LATER. The key word that applies here. About a million years or more later.
Oh, and once again, there's those three words I'm going to keep humiliating you with - lithic wear analysis. You know, the method of determining the ACTUAL use a tool was put to?
Except that, oh wait, none of the tools associated with Australopithecus have been determined to be wood working tools. What did the Nature paper announcing the discovery of those tools say? Oh, that's right, that they were FOOD PREPARATION TOOLS. Here's the details again:
From the paper in more detail:
Shortly after, the authors note this:
The authors conclude with:
Oh wait, what was that I was saying about TOOL USE FOR FOOD ACQUISITION AND PROCESSING again? A postulate that is merely reinforced by this latest discovery?
Note how the authors in that paper, consistently refer to removing flesh from bone for consumption. NOT the use of those tools to beat the shit out of big predators.
Nice try at sneaking in your preconceived conclusion, JayJay, but my bullshit radar is in perfect working order.
Except that THE AUTHORS OF THE PAPER CLEARLY STATE THAT THE TOOLS WERE USED FOR FOOD PREPARATION. Is this going to sink in eventually, or do I have to etch the fucking words on your forehead with a Borazon cutting awl?
Except that once again, THE AUTHORS OF THE PAPER CLEARLY STATE THAT THE TOOLS WERE USED FOR FOOD PREPARATION. Drop the fucking duplicity JayJay, it's becoming tiresome, and only serves to make a spectacle of you in a globally accessible public arena.
The answer is simple, JayJay, instead of engaging in synthetic bleating and whingeing every time you're caught out engaging in discoursive mendacity, stop posting lies.
Why the fuck should I be in the least bit civil, to someone who has repeatedly posted lies about my contributions here?
My concept of proper conduct of discourse involves not posting manifest lies. Going to learn this elementary concept sometime?
Oh, and what level of "expertise" are we talking about here? If this individual's level of "expertise" matches yours, I'm not going to be in the least bit impressed.
Oh, taken to quote mining my fucking post, have you? An entirely predictable piece of duplicity, especially as I provided far more detail in the post in question, which you predictably have duplicitously snipped out. Apparently even my providing a detailed explanation of the requisite dental morphology terms didn't stop you. And you wonder why people here in this thread universally regard you with utter scorn, derision and contempt?
No they don't. What part of "the canines of Sahelanthropus were no larger than 11 mm in the apicocoronal axis" do you not understand? Also, what part of "absence of of a honing C-P3 complex (referred to also in the Lovejoy paper as the absence of the sectorial canine complex) do you not understand? Which is now reported to be the case for both Ardipithecus AND the earlier Sahelanthropus?
Oh, so you're going to ignore the contents of the primary scientific paper I brought here, and instead bring in a Wikipedia photo you can make shit up about? Does your mendacity know no bounds?
Translation: "All DATA not conforming to my fantasies can be ignored" ... more predictable duplicity.
Except that once again, oh dear, those same small canines appear in taxa pre-dating Australopithecus by as much as 3 million years.
Shows how little attention you pay to anything not genuflecting before your fantasies, doesn't it?
Oh look, another specious attempt to hand-wave away DATA that's inconvenient for your fantasies. What part of the words "empirical test to determine wear patterns" did you miss the first time they were brought here?
Once again, JayJay, the whole fucking point, which you missed by several light years, is that scientists are diligently working to determine proper, rigorous methods for the determination of the nature of the physical interactions the requisite entities have taken part in. Indeed, this is part of the entire basis of science, as those of us who paid attention in class know only too well. Instead of simply making shit up, and pretending that said made up shit dictates how reality behaves, as you and every other creationist does, scientists are asking themselves the elementary question "what DATA would allow us to differentiate between the classes of interaction of interest we are studying?" As a consequence of so doing, and determining proper procedures for specifying the data points that would be members of the requisite sets, they're building the databases required that will allow them to determine what usages stone tooles were actually put to, as opposed to fantasising about this and pretending that real living organisms conformed to those fantasies. That's how Dominguez-Rodrigo et al were able to provide unambiguous evidence that Acheulian stone tools dating back 1.7 million years before present were used as woodworking tools, and how McPherron et al were able to determine that the 3.3 million year old tools found at Dikika were food processing tools, because they knew what DATA to look for. Erecting specious apologetic elisions to try and avoid these facts merely makes your apologetics look even more duplicitous than they already are.
Lie.
Oh wait, this is another of those elementary concepts you appear to have missed in the requisite classes. Namely, that scientists only concentrate upon a particular subset of interactions of interest, when there exist reasons for them to do so. Until then, they operate on the basis that all interactions in the set may be possible, and then set about looking for the DATA that will allow them to differentiate between the different interactions, and narrow down the possible options. Your above attempt to misrepresent the conduct of the scientists in question is even more duplicitous, given that I've already given you a thorough schooling with respect to your mendacious attempt to quote mine the Treves & Palmqvist paper, which presented vast quantities of DATA you ignored during your sleazy quote mining exercise, in order to try and prop up your data-free, evidence-free fantasies. No one is fooled by your discoursive shell games in this thread any more, JayJay, you've been exposed as a repeat offender with respect to peddling blatant falsehoods and fabrications, along with specious accusations of scientific misconduct on the part of people with genuine knowledge in the field, whose toenail clippings possess more integrity than is evident in your posts.
Ah it's standard "creationist excuse" time again. Except that YOU are the one peddling the assertion, that beating the shit out of predators was purportedly the SOLE reason for Australopithecines engaging in tool use, which means that NO other usage marks arising from other usages should, according to your fantasy narrative, ever appear on those tools. YOU are the one erecting the assertion that beating the shit out of predators purportedly constituted the sole overarching reason for a whole raft of Australopithecine behaviours and morphology, to the complete exclusion of such elementary matters as food, sex, and social interaction, all of which you continue to ignore as being part of the lives of these organisms, whilst pursuing your sad little obsessive-compulsive ideological masturbation fantasy. Your performance here is pathetic in the extreme, and there are primary school children I know of who would point and laugh at the discoursive spectacle you present. Since YOU continue to assert that beating the shit out of predators, was purportedly the sole overarching activity governing Australopithecine behaviour and morphology, it follows as a logical corollary of your asserting this, that the ONLY wear marks that would appear on their tools, if your assertion was something other than a fantasy, would be wear marks associated with weapon use, and NO other usage marks would ever appear.
But of course, you were doubtless hoping no one would see through your smoke and mirrors exercise in this respect.
Ha ha ha ha ha!
So asserts the pedlar of repeatedly exposed lies and fabrications. Except that once again, the thread provides a wealth of DATA destroying your assertions.
Poppycock. This really is fucking desperate on your part JayJay. Oh wait, what was it McPherron et al stated in their Nature paper on the Dikika fossils? Oh that's right, they stated this:
A little later on, the authors provide this:
Oh look. It's not just the tools that bear markings of usage wear, it's the substrates that those tools are used upon as well.
A little later on, the authors provide this:
The authors move on thus:
So much for your fantasy about Australopithecines forming their own military-industrial complex.
Indeed, your fatuous and desperate assertion about weapon usage purportedly leaving no marks. is such a blatant piece of evasion, that it's risible. Not least because your assertion directly contradicts everything that is taught in forensic science classes about transference. Namely, the principle that any two materials in mechanical contact will leave upon each other traces of that contact. This covers everything from surface skin molecules being left upon a touched surface (heard of fingerprint analysis, have you?) through to abrasion marks left on bullets as they are shot out of a gun. Indeed, palaeontologists press into service frequently the fact that these mechanical contact marks are persistent, sufficiently so for them to be ecologically informative with respect to 80 million year old fossils. Which, funnily enough, I schooled you on earlier in this post with respect to the determination of the ecological niche of Tyrannosaurus rex.
I've noticed how creationists don't bother with joined up thinking.
I just did. By covering everything from the actual scientific analysis of the materials in question, through to fisking your preposterous assertion that contradicts everything taught in forensic science classes. This is how it's done properly, JayJay, not that you'll ever learn from this.
Well here we're into the territory of your lack of rigour once more. Because first of all, a defensive weapon in an ecosystem is at its most effective, when it provides the assailant being fended off with enough pain to think twice about repeating the assault, whilst leaving said assailant without permanent or life threatening injury. A dead predator can't pass on lessons about the dangers of certain would-be prey either to its companions or its offspring. In order for defensive weapons to be effective, they not only have to protect the possessor, but pass on a persistent message to would-be assailants, that an attack upon said possessors is a bad idea. Which, by definition, can only happen if assailants survive such encounters, remember said encounters, and communicate the information about this to other members of their species.
This then brings us on to the matter of how much force is required to deliver this result. But of course, even a relatively modest force can still result in persistent transference, that can be searched for in the requisite items, be those items the tools used for defence, or the fossilised skeletal remains of the assailant. Now since scientists know what sort of percussion marks to look for, half of the jigsaw is complete if they find fossils of predators bearing stone tool abrasion marks. Once we have that in place, we're on the road to having DATA supporting the hypothesis.
Now, last time I checked my basic ecology classes, in consilience with my above remarks about when a defensive weapon is truly effective, most armed organisms don't set out to kill their predators. They set out simply to fend off those predators, ideally implanting a persistent memory to the effect that the defenders are a long way removed from being easy meat. The best modus operandi for defence in an ecosystem isn't to inflict grave injuries upon your assailant, as I stated above, but merely to make the attack memorably painful and fruitless for the attacker. Indeed, that's one elementary lesson of conflict that quite a few humans forget.
Consequently, the question genuinely requiring answering is not "what characteristics would make a defensive weapon lethal or critically injurious", but "what characteristics would make a defensive weapon deliver the persistent deterrent message most effectively?" Which renders all your talk about "crushing skulls" naively infantile when considering how real ecosystem interactions operate.
Now of course, whilst sharp implements require less force to deploy in order to be painful than blunt implements, they require somewhat more precise control, to deploy in a manner that doesn't exceed the requirements of successful deterrence. In the heat of the moment, a defender could easily, in desperation, use more force than necessary for deterrence, though it might not appear that way to the defender during the attack. Consequently, the ideal defensive weapon is one that doesn't require exquisite motor skills to deploy, so that it's easy to use effectively when caught by surprise, and doesn't grossly exceed the minimum deterrence requirement. A hand axe, being a generalist tool, would be a reasonable candidate for an organism already familiar with the usage thereof for other purposes, but this doesn't mean for one moment that actual hominid hand axes were used in that manner. The only time we can conclude this, is when we have proper evidence for such usage, so my answer doesn't help your apologetics in the slightest, and I'll be watching assiduously for signs of you quote mining this paragraph of mine in future responses.
So fucking what? I was simply pointing out the absurdity of your assertion about the purported perishability of wood. Or did you not notice this in your desperation to fire off another cheap apologetic rejoinder?
Except that, oh wait, once again, we have NO evidence that Australopithecines engaged in any of this behaviour you're attributing thereto. The ONLY evidence for tool use amongst Australopithecines to date, centres upon food preparation. When is this elementary fact going to sink in?
Pull the other one, it's got fucking bells on!
You've been peddling a manifest fantasy about Australopithecines behaving in precisely this manner. My satirical rendition of your fantasy is stinging you, because it's too uncomfortably close to the nebulous hologram you've cooked up in the television inside your head. You're busted, JayJay, just as you were busted some years back over your aquarium ornament débacle.
I haven't seen many girls with brow ridges of late ...
Bollocks. Go and read all those scientific papers I've presented, and learn why their contents destroy your fantasy. And by "read", I don't mean "skim for quote mines".
It's as far removed from the truth as right wing politics.
Bullshit. How many times have you modified your behaviour to obtain sex, JayJay? Don't fucking lie here, every male human on the fucking planet has done this at some point in his life. He's changed his behaviour to make him more desirable to the woman he's hoping to shag. That you are completely oblivious to this, makes me wonder if you've ever had fucking sex. Fucking hell, JayJay, there are thirteen year olds that know better than you on this one.
Jayjay4547 wrote:Calilasseia wrote: 2: ALL tools reliably associated with Australopithecus have been FOOD PREPARATION TOOLS;
One of the uses of an Oldowan hand axe would have been to ding the skull of a predator. Edged tools have been used for woodworking as you say later in your post- that isn’t food preparation.
LATER. The key word that applies here. About a million years or more later.
Oh, and once again, there's those three words I'm going to keep humiliating you with - lithic wear analysis. You know, the method of determining the ACTUAL use a tool was put to?
Jayjay4547 wrote:What do you imagine Australopithecus might have whittled away at wood FOR?
Except that, oh wait, none of the tools associated with Australopithecus have been determined to be wood working tools. What did the Nature paper announcing the discovery of those tools say? Oh, that's right, that they were FOOD PREPARATION TOOLS. Here's the details again:
McPherron et al, 2010 wrote:The oldest direct evidence of stone tool manufacture comes from Gona (Ethiopia) and dates to between 2.6 and 2.5 million years (Myr) ago1. At the nearby Bouri site several cut-marked bones also show stone tool use approximately 2.5 Myr ago2. Here we report stone-tool-inflicted marks on bones found during recent survey work in Dikika, Ethiopia, a research area close to Gona and Bouri. On the basis of low-power microscopic and environmental scanning electron microscope observations, these bones show unambiguous stone-tool cut marks for flesh removal and percussion marks for marrow access. The bones derive from the Sidi Hakoma Member of the Hadar Formation. Established 40Ar–39Ar dates on the tuffs that bracket this member constrain the finds to between 3.42 and 3.24 Myr ago, and stratigraphic scaling between these units and other geological evidence indicate that they are older than 3.39 Myr ago. Our discovery extends by approximately 800,000 years the antiquity of stone tools and of stone-tool-assisted consumption of ungulates by hominins; furthermore, this behaviour can now be attributed to Australopithecus afarensis.
From the paper in more detail:
McPherron et al, 2014 wrote:DIK-55-2 (Fig. 2 and Supplementary Information) is a right rib fragment of a large ungulate, probably size 4 (cow-sized) or larger. Marks A1 and A2 are perpendicular to the cortical surface, V-shaped in cross-section with internal microstriations and diagnosed as high-confidence stone-tool cut marks. Mark B is a more obliquely oriented mark that shaves off the bone surface within which are microstriations, all consistent with a high-confidence stone-tool-inflicted mark from cutting, scraping and/or percussion. An indentation (mark C) with microstriations and crushing of the bone surface is a high-confidence hammerstone percussion mark described in Supplementary Information.
DIK-55-3 (Fig. 3 and Supplementary Information) is a femur shaft fragment of a size 2 (goat-sized) young bovid. The surface is densely marked (Fig. 3a). Mark A is perpendicular to the cortical surface and has clear microstriations running out one end (Fig. 3b, c), diagnosed as a high-confidence cut mark. Mark D (Fig. 3d–f) is a dense cluster. One prominent mark within D (Fig. 3d) has crushing of the bone surface with microstriations and is diagnosed as a high-confidence percussion mark. Mark E (Fig. 3g, h) is obliquely oriented, shaves off surface bone, has microstriations and a shouldered edge highly consistent with a stone-tool cut mark. Marks H1 and H2 overlap. H1 has clear microstriations, is associated with the broken edge of the bone and swirls in a way typical of a percussion mark. H2 shaves off bone surface and has clear microstriations, consistent with stone-tool cut marks and a scraping motion. DIK-55-3 has other high-confidence stone-tool-inflicted marks, and there is at least one mark (mark I) of uncertain agency (Supplementary Information). This specimen does not have any notches of the type that are sometimes associated with hammerstone percussion on long bones8–10, but this may be owing to post-depositional breakage of the edges that removed such notches.
The cut marks demonstrate hominin use of sharp-edged stone to remove flesh from the femur and rib. The location and density of the marks on the femur indicate that flesh was rather widely spread on the surface, although it is possible that there could have been isolated patches of flesh. The percussion marks on the femur demonstrate hominin use of a blunt stone to strike the bone, probably to gain access to the marrow. The external surfaces of ribs have thin sheaths of flesh, so the scraping marks on the fossil rib suggest stripping off of these sheaths.
Shortly after, the authors note this:
McPherron et al, 2014 wrote:It is not possible to demonstrate from the modified bones whether the stone tools were knapped for this purpose or whether naturally occurring sharp-edged stones were collected and used. No stone artefacts or sharp-edged stones were found in association with the bones at DIK-55. However, stone tool production and consequently archaeological accumulations are not expected at this locality given the sedimentary environment characterized by the palaeo-Awash River emptying into a nearby lake3,4. In this relatively low-energy depositional environment, clasts suitable for stone tool production are not present (few particles larger than fine gravel, 8 mm diameter). Within the exposed SH Member, the distance from DIK-55 to cobble-sized raw materials (>64 mm) is ~6 km (at Gorgore; Fig. 1). Thus, in this instance the absence of evidence for stone tool production in the immediate vicinity of the cut-marked bones may reflect landscape-level raw material constraints.
The authors conclude with:
McPherron et al, 2014 wrote:The bones presented here are the earliest evidence for meat and marrow consumption in the hominin lineage, pre-dating the known evidence by over 800 kyr2. Pending new discoveries, the only hominin species present in the Lower Awash Valley at 3.39 Myr ago to which we can associate this tool use is A. afarensis[i]5,15. Whether [i]A. afarensis also produced stone tools remains to be demonstrated, but the DIK-55 finds may fit with the view that stone tool production predates the earliest known archaeological sites and was initially of low intensity (one-to-a-few flakes removed per nodule) and distributed in extremely low density scatters across the landscape such that its archaeological visibility is quite low16. The evidence presented here offers a first insight into an early phase of stone tool use in hominin evolution that will improve our understanding of how this type of behaviour originated and developed into later, well recognized, stone tool production technologies.
Oh wait, what was that I was saying about TOOL USE FOR FOOD ACQUISITION AND PROCESSING again? A postulate that is merely reinforced by this latest discovery?
Note how the authors in that paper, consistently refer to removing flesh from bone for consumption. NOT the use of those tools to beat the shit out of big predators.
Nice try at sneaking in your preconceived conclusion, JayJay, but my bullshit radar is in perfect working order.
Jayjay4547 wrote:The most plausible task would have been, to fashion a pointed stick to keep predators at arms length.
Except that THE AUTHORS OF THE PAPER CLEARLY STATE THAT THE TOOLS WERE USED FOR FOOD PREPARATION. Is this going to sink in eventually, or do I have to etch the fucking words on your forehead with a Borazon cutting awl?
Jayjay4547 wrote:Sure as hang, if you slept in a cave and a leopard appeared at the entrance, who had already eaten your aunt and two nephews, and you could whittle away at wood, you would make a pointy stick.
Except that once again, THE AUTHORS OF THE PAPER CLEARLY STATE THAT THE TOOLS WERE USED FOR FOOD PREPARATION. Drop the fucking duplicity JayJay, it's becoming tiresome, and only serves to make a spectacle of you in a globally accessible public arena.
Jayjay4547 wrote:Calilasseia wrote:Jayjay4547 wrote:The other day I asked a local expert on primates “Do primates bite?” and she said yes, and that her body is full of scars as evidence of that. When I looked tremendously interested she thought it was her body that excited my interest and became a bit stiffly cold.
If your approaches to her were the same as your approaches to us with respect to the proper conduct of discourse, I'm not in the least surprised about that last part.
My approaches to posters here doesn’t include calling anyone+- a liar, a masturbatory fantasist, talking bullshit, etc etc.
The answer is simple, JayJay, instead of engaging in synthetic bleating and whingeing every time you're caught out engaging in discoursive mendacity, stop posting lies.
Jayjay4547 wrote:Your concept of “proper conduct of discourse” involves zero commitment to civility.
Why the fuck should I be in the least bit civil, to someone who has repeatedly posted lies about my contributions here?
My concept of proper conduct of discourse involves not posting manifest lies. Going to learn this elementary concept sometime?
Jayjay4547 wrote:Besides that, you sidestep that expert’s account did support the view that other primates are proficient and ready biters.
Oh, and what level of "expertise" are we talking about here? If this individual's level of "expertise" matches yours, I'm not going to be in the least bit impressed.
Jayjay4547 wrote:.Calilasseia wrote:Jayjay4547 wrote:Here is a pic of Sahelanthropus showing that it had longer canines than Australopithecus , suggesting that the adaptation was in progress roughly around that time.
A New Hominid From The Upper Miocene Of Chad, Central Africa by Michel Brunet, and others here]Brunet et al, 2002 wrote:Differential diagnosis. Sahelanthropus is distinct from all living great apes in the following respects: relatively smaller canines with apical wear, the lower showing a full occlusion above the well-developed distal tubercle, probably correlated with a non-honing C–P3 complex (P3 still unknown).Brunet et al, 2002 wrote:Sahelanthropus is distinguished as a hominid from large living and known fossil hominoid genera in the following respects: …from Gorilla by smaller canines and lower-cusped cheek teeth
]from Ouranopithecus…by markedly smaller but mesiodistally longer canines, apical wear and large distal tubercle in lower canines
Frim Sivapithecus16, smaller canines with apical wear, and thinner cheek-teeth enamel
Sahelanthropus is also distinct from all known hominid genera in the following respects:
from Homo ….non-incisiform and mesiodistally long canines,
from Ardipithecus[/i….; less incisiform upper canines not diamond shaped with a low distal shoulder and a mesiodistal long axis, bucco-lingually narrower lower canines with stronger distal tubercle, and P4 with two roots;….
from [i]Orrorin … non-chimp-like upper canines with extensive apical wear.
Again, note the frequent references to small canines with apical wear
I extracted from your quote above, the bits directly relating to canines.
Oh, taken to quote mining my fucking post, have you? An entirely predictable piece of duplicity, especially as I provided far more detail in the post in question, which you predictably have duplicitously snipped out. Apparently even my providing a detailed explanation of the requisite dental morphology terms didn't stop you. And you wonder why people here in this thread universally regard you with utter scorn, derision and contempt?
Jayjay4547 wrote:They do support my claim that this possible ancestors of Australopithecus had canines intermediate between those of apes and Australopithecus.
No they don't. What part of "the canines of Sahelanthropus were no larger than 11 mm in the apicocoronal axis" do you not understand? Also, what part of "absence of of a honing C-P3 complex (referred to also in the Lovejoy paper as the absence of the sectorial canine complex) do you not understand? Which is now reported to be the case for both Ardipithecus AND the earlier Sahelanthropus?
Jayjay4547 wrote:Here’s a pic of another possible ancestor Ardipithecus Ramidus- without canines that show “extensive apical wear” (ie worn down from on top)
Oh, so you're going to ignore the contents of the primary scientific paper I brought here, and instead bring in a Wikipedia photo you can make shit up about? Does your mendacity know no bounds?
Jayjay4547 wrote:It’s not essential to pin down the habits of ancestors of Australopithecus
Translation: "All DATA not conforming to my fantasies can be ignored" ... more predictable duplicity.
Jayjay4547 wrote:to see that even the early Australopithecus afarensis canines shows that they had already fully responded to relief from having to bite their predators.
Except that once again, oh dear, those same small canines appear in taxa pre-dating Australopithecus by as much as 3 million years.
Jayjay4547 wrote:Calilasseia wrote:Shea, 1988 wrote:Lithic use-wear analysis can provide functional infomation about stone tool assemblages and insights into prehistoric hominid behaviour. The criteria by which hafter stone spear- and arrow-points can be recognised have been extensively documented by other researchers (Ahler 1971; Fischer, Hansen and Rasmussen 1984; Odell and Cowan 1986). Indentifying such wear patterns in the Middle Palaeolithic can indicate both the geographic and temporal variation of prehistoric hunting strategies. Wear patterns on stone tools from the Levantine Middle Palaeolithic sites of Kebara, Qazfeh, and Hayonim caves suggest that technologically-assisted hunting may have been an important part of hominid subsistence as early as 50,000-100,000 years ago.
Oh wait, what did I spend several posts in this thread schooling you on? LITHIC WEAR ANALYSIS. Namely, the analysis of the different tool usage marks that different tool usages leave upon the tools in question, and which scientists have been pressing into service to analyse ACTUAL tool usage. Yet more DATA you keep ignoring. Indeed, I even schooled you on the fact that scientists went to the trouble of making their own prehistoric stone tools, in accordance with the known toolmaking methods deployed by earlier hominids, and then subjected those tools to a range of usages, to provide a database of use-wear analysis data. Indeed, I told you about this in this same post in which I provided the paper discussing the use of this technique to determine actual tool usage. I cited no less than four peer reviewed scientific works covering this topic in that post, and presented the full papers in two of those instances. So your assertion that I didn't back up my statement is a BARE FACED LIE.
This is all you have to offer here, JayJay, made up shit accompanied by bare faced lies and other well-documented forms of creationist duplicity. I and others here, on the other hand, have REAL DATA to bring to the table.
Now, since I've already made numerous references to lithic wear analysis, and how this allows scientists to distinguish which usage a given tool was put to, I think we can all safely conclude that I've provided more than enough substance allowing a weapon to be distinguished from, say, a food preparation or materials handling tool, because, lo and behold, as scientists have discovered experimentally, all of these usages leave distinct and analysable wear patterns on the tools in question. A weapon would have different usage marks to a food preparation tool, which in turn would have different usage marks to a materials processing tool. Indeed, the Shea paper I provided earlier mentions a specific experiment conducted to establish use-wear patterns on stone tools, viz:Shea, 1988 wrote:Whatver approach or combination of approaches is employed, archaeological analyses must be accompanied by a demonstration of each analyst's ability to reliably reconstruct the functions of experimentally-utilised stone tools. It is only from such a demonstration that accuracy levels and confidence intervals can be assigned to archaeological interpretations. In the course of learning the "low magnification" approach, six independent blind tests of the author's ability to reconstruct stone tool uses were conducted over three years (Shea 1987). These tests involved a total of 243 uses on 111 artefacts (with 33 unused "dummy" specimens). All artefacts had been used, without the author's participation or observation, by graduates of lithic technology workshops and had been subjected to simulated archaeological screening and washing. The test results suggest the following accuracy levels and confidence intervals for archaeological application: location of use = 237/243 correct (x=97%±1%); action employed (of 16 possibilities) = 222/243 correct (x=91%±5%), worked material (of 12 possibilities) = 200/243 correct (x=82%±5%). No statistically significant differences were noted in the accuracy rates for different actions and worked materials, or between retouched and unretouched edges. Only the duration of tool use and the type of lithic raw material appeared to significantly affect analytical accuracy.
So the author not only engaged in an experiment aimed at determining which wear patterns accompany which tool usage, but also aimed at determining his ability to deduce correctly the requisite wear usage. Now the mere fact that I brought that paper here, on its own tells anyone appraising this thread honestly, that I did more than sufficient to establish my case. Namely, that scientists have rigorous methods allowing them to tell the difference between tools used as weapons, tools used for food preparation, and tools used for material processing by prehistoric hominids. They have the DATA. The mere fact that you couldn't be bothered to examine that paper, alight upon this DATA yourself, then fabricated a bare faced lie to the effect that I failed to support my statements robustly in this vein, again destroys your fantasy.
I did miss your citing a paper that dated the earliest weapons to 100 000 years ago, being hafted hunting weapons.
Shows how little attention you pay to anything not genuflecting before your fantasies, doesn't it?
Jayjay4547 wrote:As to lithic wear patterns, a lithic laboratory workshop in which participants didn’t face predation would be a markedly poor reconstruction of Australopithecus natural context.
Oh look, another specious attempt to hand-wave away DATA that's inconvenient for your fantasies. What part of the words "empirical test to determine wear patterns" did you miss the first time they were brought here?
Once again, JayJay, the whole fucking point, which you missed by several light years, is that scientists are diligently working to determine proper, rigorous methods for the determination of the nature of the physical interactions the requisite entities have taken part in. Indeed, this is part of the entire basis of science, as those of us who paid attention in class know only too well. Instead of simply making shit up, and pretending that said made up shit dictates how reality behaves, as you and every other creationist does, scientists are asking themselves the elementary question "what DATA would allow us to differentiate between the classes of interaction of interest we are studying?" As a consequence of so doing, and determining proper procedures for specifying the data points that would be members of the requisite sets, they're building the databases required that will allow them to determine what usages stone tooles were actually put to, as opposed to fantasising about this and pretending that real living organisms conformed to those fantasies. That's how Dominguez-Rodrigo et al were able to provide unambiguous evidence that Acheulian stone tools dating back 1.7 million years before present were used as woodworking tools, and how McPherron et al were able to determine that the 3.3 million year old tools found at Dikika were food processing tools, because they knew what DATA to look for. Erecting specious apologetic elisions to try and avoid these facts merely makes your apologetics look even more duplicitous than they already are.
Jayjay4547 wrote:So the possible use of an Oldowan hand axe to smash a predator skull wasn’t even considered.
Lie.
Oh wait, this is another of those elementary concepts you appear to have missed in the requisite classes. Namely, that scientists only concentrate upon a particular subset of interactions of interest, when there exist reasons for them to do so. Until then, they operate on the basis that all interactions in the set may be possible, and then set about looking for the DATA that will allow them to differentiate between the different interactions, and narrow down the possible options. Your above attempt to misrepresent the conduct of the scientists in question is even more duplicitous, given that I've already given you a thorough schooling with respect to your mendacious attempt to quote mine the Treves & Palmqvist paper, which presented vast quantities of DATA you ignored during your sleazy quote mining exercise, in order to try and prop up your data-free, evidence-free fantasies. No one is fooled by your discoursive shell games in this thread any more, JayJay, you've been exposed as a repeat offender with respect to peddling blatant falsehoods and fabrications, along with specious accusations of scientific misconduct on the part of people with genuine knowledge in the field, whose toenail clippings possess more integrity than is evident in your posts.
Jayjay4547 wrote:And if a hand axe was used for that purpose, it might well not have been used often enough to show wear patterns
Ah it's standard "creationist excuse" time again. Except that YOU are the one peddling the assertion, that beating the shit out of predators was purportedly the SOLE reason for Australopithecines engaging in tool use, which means that NO other usage marks arising from other usages should, according to your fantasy narrative, ever appear on those tools. YOU are the one erecting the assertion that beating the shit out of predators purportedly constituted the sole overarching reason for a whole raft of Australopithecine behaviours and morphology, to the complete exclusion of such elementary matters as food, sex, and social interaction, all of which you continue to ignore as being part of the lives of these organisms, whilst pursuing your sad little obsessive-compulsive ideological masturbation fantasy. Your performance here is pathetic in the extreme, and there are primary school children I know of who would point and laugh at the discoursive spectacle you present. Since YOU continue to assert that beating the shit out of predators, was purportedly the sole overarching activity governing Australopithecine behaviour and morphology, it follows as a logical corollary of your asserting this, that the ONLY wear marks that would appear on their tools, if your assertion was something other than a fantasy, would be wear marks associated with weapon use, and NO other usage marks would ever appear.
But of course, you were doubtless hoping no one would see through your smoke and mirrors exercise in this respect.
Jayjay4547 wrote:Calilasseia wrote:
Which, funnily enough, is followed shortly after by the entire section on evidence from wear analysis that you never bothered reading. I wonder why you never bothered reading it? would this have something to do with the fact that all the scientific literature pertinent to this DESTROYS your fantasy assertions?
The only fantasy around here is your overbearing fantasy about destroying propositions that you don’t like.
Ha ha ha ha ha!
So asserts the pedlar of repeatedly exposed lies and fabrications. Except that once again, the thread provides a wealth of DATA destroying your assertions.
Jayjay4547 wrote:Calilasseia wrote:Jayjay4547 wrote:Would a primate holding a hand axe like that, be dangerous to attack? My sense is yes but with a major reservation; something is missing; a means to keep a predator at arms length.
Oh wait, what usage wear analysis exists to support your fantasy about weapon wielding Australopithecines? NONE.
A stone like that could have crushed many a skull without showing wear patterns.
Poppycock. This really is fucking desperate on your part JayJay. Oh wait, what was it McPherron et al stated in their Nature paper on the Dikika fossils? Oh that's right, they stated this:
McPherron et al, 2010 wrote:In January 2009, the Dikika Research Project systematically collected fossils from localities just opposite the DIK-1 locality in the Andedo drainage, which predominantly exposes the Sidi Hakoma (SH) Member of the Hadar Formation (3.42–3.24 Myr ago; Fig. 1). Archaeological survey was conducted simultaneously in these same localities. In the course of this work, four fossils were identified with surface modifications which, based on field observations, resembled stone-tool cut marks7. These fossils were subsequently studied with optical and environmental scanning electron microscopy (ESEM) (see Methods and Supplementary Information). Secondary electron imaging (SEI) and energy dispersive X-ray (EDX) spectrometry data show that the marks on two of these fossils (DIK-55-2 and DIK-55-3) formed before fossilization. Optical and ESEM observations show that the marks lack the morphology indicative of trampling and biochemical marks, and that these two specimens have modifications clearly indicative of stone tool use, including cutting and percussion.
A little later on, the authors provide this:
McPherron et al, 2010 wrote:DIK-55-2 (Fig. 2 and Supplementary Information) is a right rib fragment of a large ungulate, probably size 4 (cow-sized) or larger. Marks A1 and A2 are perpendicular to the cortical surface, V-shaped in cross-section with internal microstriations and diagnosed as high-confidence stone-tool cut marks. Mark B is a more obliquely oriented mark that shaves off the bone surface within which are microstriations, all consistent with a high-confidence stone-tool-inflicted mark from cutting, scraping and/or percussion. An indentation (mark C) with microstriations and crushing of the bone surface is a high-confidence hammerstone percussion mark described in Supplementary Information.
DIK-55-3 (Fig. 3 and Supplementary Information) is a femur shaft fragment of a size 2 (goat-sized) young bovid. The surface is densely marked (Fig. 3a). Mark A is perpendicular to the cortical surface and has clear microstriations running out one end (Fig. 3b, c), diagnosed as a high-confidence cut mark. Mark D (Fig. 3d–f) is a dense cluster. One prominent mark within D (Fig. 3d) has crushing of the bone surface with microstriations and is diagnosed as a high-confidence percussion mark. Mark E (Fig. 3g, h) is obliquely oriented, shaves off surface bone, has microstriations and a shouldered edge highly consistent with a stone-tool cut mark. Marks H1 and H2 overlap. H1 has clear microstriations, is associated with the broken edge of the bone and swirls in a way typical of a percussion mark. H2 shaves off bone surface and has clear microstriations, consistent with stone-tool cut marks and a scraping motion. DIK-55-3 has other high-confidence stone-tool-inflicted marks, and there is at least one mark (mark I) of uncertain agency (Supplementary Information). This specimen does not have any notches of the type that are sometimes associated with hammerstone percussion on long bones8–10, but this may be owing to post-depositional breakage of the edges that removed such notches.
The cut marks demonstrate hominin use of sharp-edged stone to remove flesh from the femur and rib. The location and density of the marks on the femur indicate that flesh was rather widely spread on the surface, although it is possible that there could have been isolated patches of flesh. The percussion marks on the femur demonstrate hominin use of a blunt stone to strike the bone, probably to gain access to the marrow. The external surfaces of ribs have thin sheaths of flesh, so the scraping marks on the fossil rib suggest stripping off of these sheaths.
Oh look. It's not just the tools that bear markings of usage wear, it's the substrates that those tools are used upon as well.
A little later on, the authors provide this:
McPherron et al, 2010 wrote:It is not possible to demonstrate from the modified bones whether the stone tools were knapped for this purpose or whether naturally occurring sharp-edged stones were collected and used. No stone artefacts or sharp-edged stones were found in association with the bones at DIK-55. However, stone tool production and consequently archaeological accumulations are not expected at this locality given the sedimentary environment characterized by the palaeo-Awash River emptying into a nearby lake3,4. In this relatively low-energy depositional environment, clasts suitable for stone tool production are not present (few particles larger than fine gravel, 8mm diameter). Within the exposed SH Member, the distance from DIK-55 to cobble-sized raw materials (>64mm) is ~6 km (at Gorgore; Fig. 1). Thus, in this instance the absence of evidence for stone tool production in the immediate vicinity of the cut-marked bones may reflect landscape-level raw material constraints.
The authors move on thus:
McPherron et al, 2010 wrote:The bones presented here are the earliest evidence for meat and marrow consumption in the hominin lineage, pre-dating the known evidence by over 800 kyr2. Pending new discoveries, the only hominin species present in the Lower Awash Valley at 3.39 Myr ago to which we can associate this tool use is A. afarensis5,15. Whether A. afarensis also produced stone tools remains to be demonstrated, but the DIK-55 finds may fit with the view that stone tool production predates the earliest known archaeological sites and was initially of low intensity (one-to-a-few flakes removed per nodule) and distributed in extremely low density scatters across the landscape such that its archaeological visibility is quite low16. The evidence presented here offers a first insight into an early phase of stone tool use in hominin evolution that will improve our understanding of how this type of behaviour originated and developed into later, well recognized, stone tool production technologies.
So much for your fantasy about Australopithecines forming their own military-industrial complex.
Indeed, your fatuous and desperate assertion about weapon usage purportedly leaving no marks. is such a blatant piece of evasion, that it's risible. Not least because your assertion directly contradicts everything that is taught in forensic science classes about transference. Namely, the principle that any two materials in mechanical contact will leave upon each other traces of that contact. This covers everything from surface skin molecules being left upon a touched surface (heard of fingerprint analysis, have you?) through to abrasion marks left on bullets as they are shot out of a gun. Indeed, palaeontologists press into service frequently the fact that these mechanical contact marks are persistent, sufficiently so for them to be ecologically informative with respect to 80 million year old fossils. Which, funnily enough, I schooled you on earlier in this post with respect to the determination of the ecological niche of Tyrannosaurus rex.
I've noticed how creationists don't bother with joined up thinking.
Jayjay4547 wrote:Answer the question.
I just did. By covering everything from the actual scientific analysis of the materials in question, through to fisking your preposterous assertion that contradicts everything taught in forensic science classes. This is how it's done properly, JayJay, not that you'll ever learn from this.
Jayjay4547 wrote:Or let me put it another way: Can you imagine a better shape for a stone intended and held so as to crush a skull?
Well here we're into the territory of your lack of rigour once more. Because first of all, a defensive weapon in an ecosystem is at its most effective, when it provides the assailant being fended off with enough pain to think twice about repeating the assault, whilst leaving said assailant without permanent or life threatening injury. A dead predator can't pass on lessons about the dangers of certain would-be prey either to its companions or its offspring. In order for defensive weapons to be effective, they not only have to protect the possessor, but pass on a persistent message to would-be assailants, that an attack upon said possessors is a bad idea. Which, by definition, can only happen if assailants survive such encounters, remember said encounters, and communicate the information about this to other members of their species.
This then brings us on to the matter of how much force is required to deliver this result. But of course, even a relatively modest force can still result in persistent transference, that can be searched for in the requisite items, be those items the tools used for defence, or the fossilised skeletal remains of the assailant. Now since scientists know what sort of percussion marks to look for, half of the jigsaw is complete if they find fossils of predators bearing stone tool abrasion marks. Once we have that in place, we're on the road to having DATA supporting the hypothesis.
Now, last time I checked my basic ecology classes, in consilience with my above remarks about when a defensive weapon is truly effective, most armed organisms don't set out to kill their predators. They set out simply to fend off those predators, ideally implanting a persistent memory to the effect that the defenders are a long way removed from being easy meat. The best modus operandi for defence in an ecosystem isn't to inflict grave injuries upon your assailant, as I stated above, but merely to make the attack memorably painful and fruitless for the attacker. Indeed, that's one elementary lesson of conflict that quite a few humans forget.
Consequently, the question genuinely requiring answering is not "what characteristics would make a defensive weapon lethal or critically injurious", but "what characteristics would make a defensive weapon deliver the persistent deterrent message most effectively?" Which renders all your talk about "crushing skulls" naively infantile when considering how real ecosystem interactions operate.
Now of course, whilst sharp implements require less force to deploy in order to be painful than blunt implements, they require somewhat more precise control, to deploy in a manner that doesn't exceed the requirements of successful deterrence. In the heat of the moment, a defender could easily, in desperation, use more force than necessary for deterrence, though it might not appear that way to the defender during the attack. Consequently, the ideal defensive weapon is one that doesn't require exquisite motor skills to deploy, so that it's easy to use effectively when caught by surprise, and doesn't grossly exceed the minimum deterrence requirement. A hand axe, being a generalist tool, would be a reasonable candidate for an organism already familiar with the usage thereof for other purposes, but this doesn't mean for one moment that actual hominid hand axes were used in that manner. The only time we can conclude this, is when we have proper evidence for such usage, so my answer doesn't help your apologetics in the slightest, and I'll be watching assiduously for signs of you quote mining this paragraph of mine in future responses.
Jayjay4547 wrote:Calilasseia wrote:Jayjay4547 wrote:That primate would need a stopper tool either in its other hand or held in two hands by another member of the troop. Think of a dog attacking you- you would need something to stop it with and something to punish it with. But of course, sticks unlike stones are highly perishable and relatively unlikely to be found as fossils.
HA HA HA HA HA HA HA!
JayJay, we have in existence FOSSILS OF TREES DATING BACK ALL THE WAY TO THE CARBONIFEROUS. Under the right conditions, wood can fossilise, and persist for OVER 300 MILLION YEARS. The oldest known tree, Watteiza, dates back all the way to the middle Devonian, 385 million years before present. Archaeopteris is only slightly younger at 383 million years.
If you want to play that game, the oldest stones date back nearly ten times longer than your oldest fossil wood.
So fucking what? I was simply pointing out the absurdity of your assertion about the purported perishability of wood. Or did you not notice this in your desperation to fire off another cheap apologetic rejoinder?
Jayjay4547 wrote:Oldowan hand axes are preserved till today except for a chance finding their way into a grinding stream bed or glacial floor. But whatever sticks Australopithecus made using stone tools, would only be preserved under exceptional circumstances.
Except that, oh wait, once again, we have NO evidence that Australopithecines engaged in any of this behaviour you're attributing thereto. The ONLY evidence for tool use amongst Australopithecines to date, centres upon food preparation. When is this elementary fact going to sink in?
Jayjay4547 wrote:Calilasseia wrote:But of course, none of this in the least supports your sad little fantasy, about Australopithecines wielding weapons like some sort of prehistoric Call of Duty spec-ops troop. All the DATA says your fantasy never happened.
Call of Duty? That might be your fantasy, but I haven’t presented Australopithecus that way.
Pull the other one, it's got fucking bells on!
You've been peddling a manifest fantasy about Australopithecines behaving in precisely this manner. My satirical rendition of your fantasy is stinging you, because it's too uncomfortably close to the nebulous hologram you've cooked up in the television inside your head. You're busted, JayJay, just as you were busted some years back over your aquarium ornament débacle.
Jayjay4547 wrote:A weird habit of Australopithecus enabled their males to have girly skulls
I haven't seen many girls with brow ridges of late ...
Jayjay4547 wrote:and so unlike other primates, freed them from holding up evolution of the species.
Bollocks. Go and read all those scientific papers I've presented, and learn why their contents destroy your fantasy. And by "read", I don't mean "skim for quote mines".
Jayjay4547 wrote:OK that’s just a shadow of the truth
It's as far removed from the truth as right wing politics.
Jayjay4547 wrote:but it’s a lot more true than the story line in Darwin’s Descent of Man where female allure provoked their menfolk into evolving.
Bullshit. How many times have you modified your behaviour to obtain sex, JayJay? Don't fucking lie here, every male human on the fucking planet has done this at some point in his life. He's changed his behaviour to make him more desirable to the woman he's hoping to shag. That you are completely oblivious to this, makes me wonder if you've ever had fucking sex. Fucking hell, JayJay, there are thirteen year olds that know better than you on this one.
Signature temporarily on hold until I can find a reliable image host ...
-
Calilasseia - RS Donator
- Posts: 22642
- Age: 62
- Country: England
Re: How atheist ideology messed up the human origin story
#1011 ![]() by Sendraks » Dec 15, 2015 2:59 pm
by Sendraks » Dec 15, 2015 2:59 pm
Tell me JayJay by looking at this skull, what sort of meat did this creature eat?

"One of the great tragedies of mankind is that morality has been hijacked by religion." - Arthur C Clarke
"'Science doesn't know everything' - Well science knows it doesn't know everything, otherwise it'd stop" - Dara O'Brian
"'Science doesn't know everything' - Well science knows it doesn't know everything, otherwise it'd stop" - Dara O'Brian
-
Sendraks - Name: D-Money Jr
- Posts: 15260
- Age: 107
- Country: England
Re: How atheist ideology messed up the human origin story
#1012 ![]() by Jayjay4547 » Dec 17, 2015 5:28 am
by Jayjay4547 » Dec 17, 2015 5:28 am
Sendraks wrote:Jayjay4547 wrote:
It’s not a fantasy, not sad, or even little. It’s a strong INFERENCE that Australopithecus used hand held weapons to make them dangerous to attack. You used that word “fantasy” 16 times in your post.
Changing the word "fantasy" to "inference" doesn't make your argument any stronger JayJay. Whichever word you use, you're still making stuff up in an evidence free environment.
The short blunt canines of Australopithecus IS evidence. It’s very striking evidence that unlike other higher primates, Australopithecus was not a formidable biter.
Using the word “fantasy” 16 times in one post, which is what Calli did, served to make it seem that my point is a thing of air which it isn’t. It’s a strikingly clear and simple point that would appear to any rational person who places Australopithecus INSIDE the context of Africa rather than ON an acting stage. When the history of development of a science shows that the point hasn’t been taken, over a period of 90 years, then that itself becomes a significant point of interest.
Sendraks wrote:Jayjay4547 wrote: So the possible use of an Oldowan hand axe to smash a predator skull wasn’t even considered. And if a hand axe was used for that purpose, it might well not have been used often enough to show wear patterns,
Ahhh, classic theist argument. "just because you can't see the evidence doesn't mean it didn't happen/doesn't exist/isn't there."
If I happened across an Australopithecus unarmed that would be quite different from coming across one carrying a hand axe as illustrated. The first experience might make me reach for a camera, in the second case it would strike me that the creature was carrying a murder weapon. That perception would arise from its shape and my understanding of what a hand holding the stone is capable of doing. I wouldn’t need to call for a forensic analysis of the stone nor would the absence of traces of skin and bone on it convince me that the stone wasn’t a weapon.
Sendraks wrote: My inference is that Australopiths defended themselves using a form of highly sophisticated hand to hand combat, which employed their great strength to good effect. This combat form would've been, at least, as formidable as anything developed in China millions of years later.
The issue that is significant for reconstructing the ecology of Australopithecus – in other words, its interactions with things it wanted to metabolise, and those that wanted to metabolise it-or in other words, its condition INSIDE the African context- was what Australopithecus did with its predators. It wasn’t about what Australopithecus did with its fellow hominins. Skill in hand to hand combat wouldn’t have been any use against hyena.
Sendraks wrote: Of course the bones of Australopiths don't show any signs of these formidable technique being used to deter predators, but that probably only happened rarely, which explains that away and alleviates me from the burden of evidence.
It also alleviates me from the burden of rational discourse or the burden of anyone taking my views seriously.
Only a moron would seriously suggest that Australopithecus used “hand to hand combat” against predators including hyena. In your case, you make a moronic suggestion to obliquely imply that the inference of defensive weapon use is also moronic, which it isn’t.
Sendraks wrote: I feel wonderously floaty and light free of these constraints.
Like myself and my arguments suddenly lack all substance.
Why not try some straight discussion.
-
Jayjay4547 - THREAD STARTER
- Name: Jonathan
- Posts: 1482
- Country: South Africa
Re: How atheist ideology messed up the human origin story
#1013 ![]() by Thomas Eshuis » Dec 17, 2015 10:11 am
by Thomas Eshuis » Dec 17, 2015 10:11 am
Why not try some evidence and end to your incessant lies?
"Respect for personal beliefs = "I am going to tell you all what I think of YOU, but don't dare retort and tell what you think of ME because...it's my personal belief". Hmm. A bully's charter and no mistake."
-
Thomas Eshuis - Name: Thomas Eshuis
- Posts: 31091
- Age: 34
- Country: Netherlands
Re: How atheist ideology messed up the human origin story
#1014 ![]() by Sendraks » Dec 17, 2015 10:19 am
by Sendraks » Dec 17, 2015 10:19 am
Jayjay4547 wrote:
The short blunt canines of Australopithecus IS evidence.
The important thing to understand about evidence JayJay, is that it is not evidence of whatever you want to imagine or infer it might be.
Jayjay4547 wrote:It’s very striking evidence that unlike other higher primates, Australopithecus was not a formidable biter.
Uhh, what? Australopiths had strong jaws and a powerful bite.
Bite power =/= what sort of teeth you have.
Jayjay4547 wrote:
If I happened across an Australopithecus unarmed that would be quite different from coming across one carrying a hand axe as illustrated. The first experience might make me reach for a camera, in the second case it would strike me that the creature was carrying a murder weapon.
Why? Why would you leap to that conclusion? Beyond that it simply justifies your imagined use of weapons by australopiths.
Jayjay4547 wrote: Skill in hand to hand combat wouldn’t have been any use against hyena.
Well I imagine that Australopiths had a very sophisticated form of hand to hand combat, that was effective against Hyenas. You can't find any evidence that proves me wrong and, like yourself, I don't have to provide evidence that justifies my position.
Jayjay4547 wrote:Only a moron would seriously suggest that Australopithecus used “hand to hand combat” against predators including hyena. In your case, you make a moronic suggestion to obliquely imply that the inference of defensive weapon use is also moronic, which it isn’t.
1 - you haven't any evidence to prove me wrong.
2 - it is your attempt to assert weapon use by australopiths without any evidence that is moronic.
Jayjay4547 wrote:Why not try some straight discussion.
You first.
Last edited by Sendraks on Dec 17, 2015 4:57 pm, edited 1 time in total.
"One of the great tragedies of mankind is that morality has been hijacked by religion." - Arthur C Clarke
"'Science doesn't know everything' - Well science knows it doesn't know everything, otherwise it'd stop" - Dara O'Brian
"'Science doesn't know everything' - Well science knows it doesn't know everything, otherwise it'd stop" - Dara O'Brian
-
Sendraks - Name: D-Money Jr
- Posts: 15260
- Age: 107
- Country: England
Re: How atheist ideology messed up the human origin story
#1015 ![]() by monkeyboy » Dec 17, 2015 4:41 pm
by monkeyboy » Dec 17, 2015 4:41 pm
Jayjay4547 wrote:
If I happened across an Australopithecus unarmed that would be quite different from coming across one carrying a hand axe as illustrated. The first experience might make me reach for a camera, in the second case it would strike me that the creature was carrying a murder weapon. That perception would arise from its shape and my understanding of what a hand holding the stone is capable of doing. I wouldn’t need to call for a forensic analysis of the stone nor would the absence of traces of skin and bone on it convince me that the stone wasn’t a weapon.
I think a very important word is missing from this paragraph. It is "potential". An axe is a tool. It can be used violently for sure and we have examples from more recent times when the hand axe has evolved into a specifically designed tool for violence.
Just because an item could potentially have been used for something does not constitute evidence that it was used that way. Remember, all salmon are fish, yet not all fish are salmon.
I own 3 axes. Two are most definitely tools. Their use is for chopping wood. They have the potential to be murder weapons but that capability does not mean that is their purpose. The third axe is replica of a viking axe as found in York. It's a copy of a weapon but it still isn't a murder weapon, it's an ornament firmly fixed to the wall to prevent it's misuse as a weapon or even as a tool.
Your perception of what you think something is or could have been does not constitute evidence Jayjay4547. That is just evidence of assumption on your part. It don't make it so. Just like I'm no axe murderer because I own axes. I have the potential sure but only the evidence of me having used one to kill someone will change the axe's being a tool to being a murder weapon............not it's potential to be used as one.
Big word that "potential".
The Bible is full of interest. It has noble poetry in it; and some clever fables; and some blood-drenched history; and some good morals; and a wealth of obscenity; and upwards of a thousand lies.
Mark Twain
Mark Twain
-

monkeyboy - Posts: 5496
- Country: England
")
Re: How atheist ideology messed up the human origin story
#1016 ![]() by tolman » Dec 18, 2015 1:34 am
by tolman » Dec 18, 2015 1:34 am
Jayjay4547 wrote:Sendraks wrote:Jayjay4547 wrote:
It’s not a fantasy, not sad, or even little. It’s a strong INFERENCE that Australopithecus used hand held weapons to make them dangerous to attack. You used that word “fantasy” 16 times in your post.
Changing the word "fantasy" to "inference" doesn't make your argument any stronger JayJay. Whichever word you use, you're still making stuff up in an evidence free environment.
The short blunt canines of Australopithecus IS evidence. It’s very striking evidence that unlike other higher primates, Australopithecus was not a formidable biter.
According to the available evidence, to the extent that was true for Australopithecus, it appears to have also been the case for its ancestors.
I don't do sarcasm smileys, but someone as bright as you has probably figured that out already.
- tolman
- Posts: 7106
- Country: UK
Re: How atheist ideology messed up the human origin story
#1017 ![]() by Oldskeptic » Dec 18, 2015 2:02 am
by Oldskeptic » Dec 18, 2015 2:02 am
JayJay wrote:
The short blunt canines of Australopithecus IS evidence. It’s very striking evidence that unlike other higher primates, Australopithecus was not a formidable biter.
The short blunt canines of A. afarensis is evidence that they, their cousin species, their descendents, and ancestors going back to the chimpanzee homo split didn't smell or taste good. There was no need for sharp and tall canines, or tree climbing, or pointy sticks. Large predators simply avoided eating them because their smell was unappetizing and their flesh was unpalatable to all but the most desperate large predators.
There is nothing so absurd that some philosopher will not say it - Cicero.
Traditionally these are questions for philosophy, but philosophy is dead - Stephen Hawking
Traditionally these are questions for philosophy, but philosophy is dead - Stephen Hawking
-

Oldskeptic - Posts: 7395
- Age: 67
Re: How atheist ideology messed up the human origin story
#1018 ![]() by Oldskeptic » Dec 18, 2015 2:45 am
by Oldskeptic » Dec 18, 2015 2:45 am
Maybe we need a new thread about how science messed up the Jewish/Christian/Muslim human origin myth? How 'bout it JayJay, you ready to participate in that one? Or are you just a one thread wonder?
Here you go http://www.rationalskepticism.org/creat ... l#p2347783
Here you go http://www.rationalskepticism.org/creat ... l#p2347783
There is nothing so absurd that some philosopher will not say it - Cicero.
Traditionally these are questions for philosophy, but philosophy is dead - Stephen Hawking
Traditionally these are questions for philosophy, but philosophy is dead - Stephen Hawking
-
Oldskeptic - Posts: 7395
- Age: 67
Re: How atheist ideology messed up the human origin story
#1019 ![]() by Sendraks » Dec 18, 2015 11:06 am
by Sendraks » Dec 18, 2015 11:06 am
Oldskeptic wrote:
The short blunt canines of A. afarensis is evidence that they, their cousin species, their descendents, and ancestors going back to the chimpanzee homo split didn't smell or taste good. There was no need for sharp and tall canines, or tree climbing, or pointy sticks. Large predators simply avoided eating them because their smell was unappetizing and their flesh was unpalatable to all but the most desperate large predators.
We'd have known this important bit of information earlier, if it hadn't been for all those damned atheists mucking up the human origin story!
And it must be right too, because there is no evidence of deodorant from that time period.
"One of the great tragedies of mankind is that morality has been hijacked by religion." - Arthur C Clarke
"'Science doesn't know everything' - Well science knows it doesn't know everything, otherwise it'd stop" - Dara O'Brian
"'Science doesn't know everything' - Well science knows it doesn't know everything, otherwise it'd stop" - Dara O'Brian
-
Sendraks - Name: D-Money Jr
- Posts: 15260
- Age: 107
- Country: England
Re: How atheist ideology messed up the human origin story
#1020 ![]() by Fenrir » Dec 18, 2015 11:33 am
by Fenrir » Dec 18, 2015 11:33 am
Oldskeptic wrote:JayJay wrote:
The short blunt canines of Australopithecus IS evidence. It’s very striking evidence that unlike other higher primates, Australopithecus was not a formidable biter.
The short blunt canines of A. afarensis is evidence that they, their cousin species, their descendents, and ancestors going back to the chimpanzee homo split didn't smell or taste good. There was no need for sharp and tall canines, or tree climbing, or pointy sticks. Large predators simply avoided eating them because their smell was unappetizing and their flesh was unpalatable to all but the most desperate large predators.
And then we made sure of it by inventing fucking Old Spice.
Religion: it only fails when you test it.-Thunderf00t.
-

Fenrir - Posts: 4101
- Country: Australia
")
Who is online
Users viewing this topic: No registered users and 3 guests