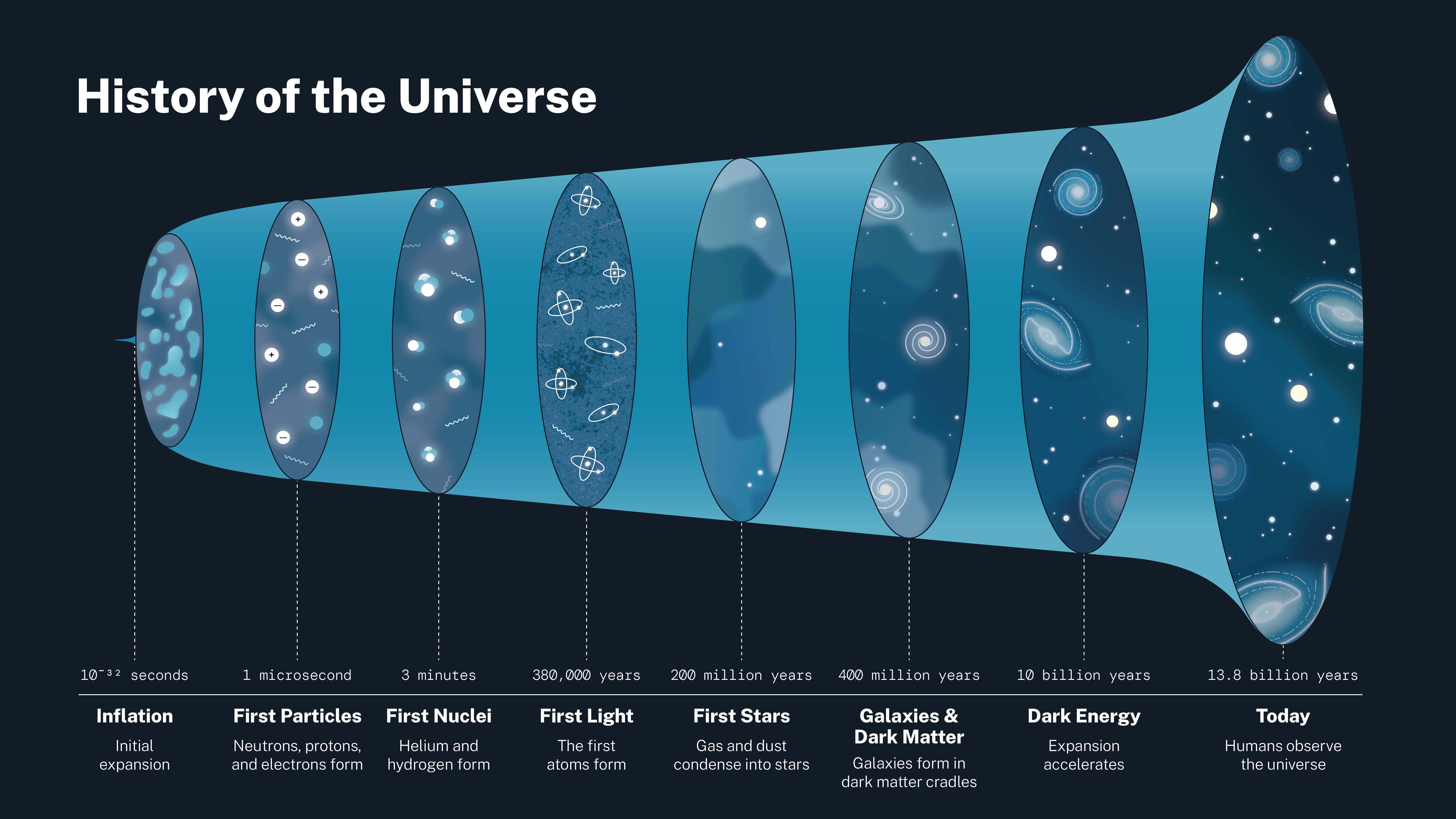
Don't mean to be pedantic, but, there's no part of this timeline that says nothing.
At best it simply doesn't show anything beyond a certain point.
interview with Alvin Plantinga
Moderators: kiore, Blip, The_Metatron
#961 ![]() by Animavore » Jun 12, 2023 6:53 pm
by Animavore » Jun 12, 2023 6:53 pm

")
#962 ![]() by Seabass » Jun 12, 2023 8:24 pm
by Seabass » Jun 12, 2023 8:24 pm
Johnny Blade wrote:My God!

#963 ![]() by Johnny Blade » Jun 13, 2023 7:56 am
by Johnny Blade » Jun 13, 2023 7:56 am
Animavore wrote:
Don't mean to be pedantic, but, there's no part of this timeline that says nothing.
At best it simply doesn't show anything beyond a certain point.
")
#964 ![]() by Spearthrower » Jun 13, 2023 9:54 am
by Spearthrower » Jun 13, 2023 9:54 am
If a human disagrees with you, let him live. In a hundred billion galaxies, you will not find another.

#965 ![]() by Rumraket » Jun 13, 2023 9:39 pm
by Rumraket » Jun 13, 2023 9:39 pm
Johnny Blade wrote:Yes. Atheism can be seen to be irrational and when used as foundation to one's worldview it leads to the belief in other absurdities and possibly even mental illness.
Johnny Blade wrote:
Here are just a few observations I believe show evidence of irrationality:
Atheists believe higher biological functions such as eye sight and consciousness arose naturally from a theoretical point in time where nothing existed except rocks.
Johnny Blade wrote:
Also I would encourage anyone who is interested to search for themselves the symptoms of anti-social and paranoid personality disorders.
#966 ![]() by THWOTH » Jun 13, 2023 10:55 pm
by THWOTH » Jun 13, 2023 10:55 pm
Johnny Blade wrote:...
Q1: Do you believe you believe there was a point in time where nothing existed in this universe except dust and rocks? (or nothing except stars..whatever you think came first, dust or stars)
Q2: Do you believe higher functions such as sight and conciousness arose naturally in the universe?

")
#967 ![]() by Spearthrower » Jun 14, 2023 3:43 am
by Spearthrower » Jun 14, 2023 3:43 am
#968 ![]() by mindhack » Jun 14, 2023 1:39 pm
by mindhack » Jun 14, 2023 1:39 pm
")
#969 ![]() by Cito di Pense » Jun 14, 2023 7:05 pm
by Cito di Pense » Jun 14, 2023 7:05 pm
Spearthrower wrote:Aye, like dusts and rocks are entities so simple as to be conceivably existent absent all the processes and forces that actually would be required to produce dust and rocks.

")
#970 ![]() by Calilasseia » Jun 15, 2023 8:38 am
by Calilasseia » Jun 15, 2023 8:38 am
Johnny Blade wrote:Yes. Atheism can be seen to be irrational and when used as foundation to one's worldview it leads to the belief in other absurdities and possibly even mental illness. Here are just a few observations I believe show evidence of irrationality:
Johnny Blade wrote:
Atheists believe higher biological functions such as eye sight and consciousness arose naturally from a theoretical point in time where nothing existed except rocks.
Pastuzyn et al (2018) wrote:
SUMMARY
The neuronal gene Arc is essential for long-lasting information storage in the mammalian brain, mediates various forms of synaptic plasticity, and has been implicated in neurodevelopmental disorders. However, little is known about Arc’s molecular function and evolutionary origins. Here, we show that Arc self-assembles into virus-like capsids that encapsulate RNA. Endogenous Arc protein is released from neurons in extracellular vesicles that mediate the transfer of Arc mRNA into new target cells, where it can undergo activity-dependent translation. Purified Arc capsids are endocytosed and are able to transfer Arc mRNA into the cytoplasm of neurons. These results show that Arc exhibits similar molecular properties to retroviral Gag proteins. Evolutionary analysis indicates that Arc is derived from a vertebrate lineage of Ty3/gypsy retrotransposons, which are also ancestors to retroviruses. These findings suggest that Gag retroelements have been repurposed during evolution to mediate intercellular communication in the nervous system.
Pastuzyn et al (2018) wrote:
INTRODUCTION
Brains have evolved to process and store information from the outside world through synaptic connections between interconnected networks of neurons. Despite the fundamental importance of information storage in the brain, we still lack a detailed molecular and cellular understanding of the processes involved and their evolutionary origins. Eukaryotic genomes are littered with DNA of viral or transposon origin, which compose about half of most mammalian genomes (Smit, 1999). A growing body of evidence indicates the sequences encoded by these elements can provide raw material for the emergence of new functions and regulatory elements (Chuong et al., 2017). In vertebrates, these include dozens of protein-coding genes derived from sequences previously encoded by transposons (Feschotte and Pritham, 2007) or retroviruses (Kaneko-Ishino and Ishino, 2012). Interestingly, many of these transposon-derived genes are expressed in the brain, but their molecular functions remain to be elucidated.
The neuronal gene Arc contains structural elements foundwithin viral Group-specific antigen (Gag) polyproteins that may have originated from the Ty3/gypsy retrotransposon family (Campillos et al., 2006; Shepherd, 2017; Zhang et al., 2015), although the role these Gag elements play in Arc function has not been explored. Arc is a master regulator of synaptic plasticity in mammals and is required for protein synthesis-dependent forms of long-term potentiation (LTP) and depression (LTD) (Bramham et al., 2010; Shepherd and Bear, 2011). Arc can regulate synaptic plasticity through the trafficking of AMPA-type glutamate receptors (AMPARs) via the endocytic machinery (Chowdhury et al., 2006). This endocytic pathway maintains levels of surface AMPARs in response to chronic changes in neuronal activity through synaptic scaling, thus contributing to neuronal homeostasis (Shepherd et al., 2006). Arc’s expression in the brain is highly dynamic; its transcription is tightly coupled to encoding of information in neuronal circuits in vivo (Guzowski et al., 1999). Arc mRNA is transported to dendrites and becomes enriched at sites of local synaptic activity where it is locally translated into protein (Steward et al., 1998; Waung et al., 2008). Intriguingly, aspects of Arc mRNA regulation resemble some viral RNAs, as Arc contains an internal ribosomal entry site (IRES) that allows cap-independent translation (Pinkstaff et al., 2001). Arc is required in vivo to transduce experience into long-lasting changes in visual cortex plasticity (McCurry et al., 2010) and for long-term memory (Guzowski et al., 2000; Plath et al., 2006). In addition, Arc has been implicated in various neurological disorders that include Alzheimer’s disease (AD) (Wu et al., 2011), neurodevelopmental disorders, such as Angelman (Greer et al., 2010; Pastuzyn and Shepherd, 2017) and Fragile X syndrome (Park et al., 2008), and schizophrenia (Fromer et al., 2014; Manago` et al., 2016; Purcell et al., 2014). Thus, precise regulation of Arc expression and activity in the nervous system seems essential for normal cognition.
Despite its importance, little is known about Arc protein biochemistry and molecular function. Here, we uncover a potential role for Arc in mediating intercellular communication via extracellular vesicles (EVs). Synaptic communication is modulated by many other communication pathways that include glia-neuron interactions, and emerging evidence suggests that EVs mediate intercellular signaling in the nervous system (Budnik et al., 2016; Zappulli et al., 2016). EVs can be broadly divided into two groups, microvesicles and exosomes, which are defined both by size and subcellular origin. Microvesicles pinch off from the plasma membrane directly and are usually 100– 300 nm in diameter, whereas exosomes are derived from intraluminal vesicles that originate from multivesicular bodies (MVBs) and are usually <100 nm in size. EVs can transport cargo that do not readily cross the plasma membrane, such as membrane proteins and various forms of RNA. The observation that EVs can function in the intercellular transport of these molecules within the nervous system opens an entirely new perspective on intercellular communication in the brain.
Here, we find that Arc protein self-assembles into oligomers that resemble virus capsids and exhibit several other biochemical properties seen in retroviral Gag proteins such as RNA binding. Moreover, Arc is released from neurons in EVs and is able to transfer its own mRNA into neurons. The Drosophila Arc homolog, dArc1, also forms capsids and mediates intercellular transfer of its own mRNA at the fly neuromuscular junction (Ashley et al., 2018, this issue of Cell), despite originating from a distinct retrotransposon lineage. These data suggest that co-option of retroviral-like Gag elements may have provided an evolutionary pathway for novel mechanisms that mediate intercellular signaling and have been intricately involved in the evolution of synaptic plasticity and animal cognition.
Pastuzyn et al (2018) wrote:
DISCUSSION
Here, we show that mammalian Arc protein exhibits many hallmarks of Gag proteins encoded by retroviruses and retrotransposons: self-assembly into capsids, RNA encapsulation, release in EVs, and intercellular transmission of RNA. These data suggest that Arc can mediate intercellular trafficking of mRNA via Arc EVs (which we term ‘‘ACBARs’’ for ‘‘Arc Capsids Bearing Any RNA’’), revealing a novel molecular mechanism by which genetic information may be transferred between neurons.
Arc Functions as a Repurposed Gag Protein
Our data show a remarkable conservation of viral Gag properties in Arc. Since Arc shows structural homology to the Gag CA domain (Zhang et al., 2015), the capability of self-assembly into oligomeric capsids is perhaps not too surprising. However, Arc seems to retain other important biochemical properties of Gag that are not intuitive from its sequence. Despite lacking clear zinc-finger RNA binding domains such as in HIV Gag, Arc encapsulates RNA, and RNA binding seems critical for capsid formation. This is reminiscent of Foamy Virus Gags, which have evolved different RNA-binding motifs to HIV Gag (Hamann and Lindemann, 2016) and also structurally resemble Arc (Taylor et al., 2017). HIV Gag-RNA interactions are complex and involve multiple components of Gag, including the MA domain, and are regulated by host cellular factors (Mailler et al., 2016). Gag MARNA interactions are also critical for virus particle formation at membranes (Kutluay et al., 2014). Moreover, if viral RNA is not present, Gag encapsulates host RNA, and any single-stranded nucleic acid longer than ~20–30 nt can support capsid assembly (Campbell and Rein, 1999), indicating a general propensity to bind abundant RNA. Indeed, precisely how viral RNA is preferentially packaged into Gag capsids in cells remains an intensive area of investigation (Comas-Garcia et al., 2016).
Pastuzyn et al (2018) wrote:
Evolution of Synaptic Plasticity and Cognition
Ty3/gypsy retrotransposons are ancient mobile elements that are widely distributed and often abundant in eukaryotic genomes and are considered ancestral to modern retroviruses (Malik et al., 2000). There is evidence that coding sequences derived from Ty3/gypsy and other retroviral-like elements have been repurposed for cellular functions repeatedly during evolution (Feschotte and Gilbert, 2012). For instance, multiple envelope genes of retroviral origins have been co-opted during mammalian evolution to promote cell-cell fusion and syncytiotrophoblast formation in the developing placenta (Cornelis et al., 2015). There are more than one hundred Gag-derived genes in the human genome alone (Campillos et al., 2006), and genetic KOs of their murine orthologs have revealed that some, like Arc, are essential for cognition (Irie et al., 2015). However, the molecular function of these Gag-derived proteins has been poorly characterized, and whether they were co-opted to serve similar cellular processes remains an open question. This study and the accompanying article from Ashley et al. (2018) now reveal that two distantly related Gag-derived genes have been independently co-opted in fly and tetrapod ancestors to participate in a similar process of EV-dependent intercellular trafficking of RNA in the nervous system. Given the plethora of retroelements populating eukaryotic genomes, we speculate that many other Gag proteins have been repurposed for cellular processes that await discovery in a variety of organisms.
Johnny Blade wrote:
Also I would encourage anyone who is interested to search for themselves the symptoms of anti-social and paranoid personality disorders.

#971 ![]() by pfrankinstein » Aug 15, 2023 4:48 pm
by pfrankinstein » Aug 15, 2023 4:48 pm
#972 ![]() by Spearthrower » Aug 15, 2023 5:04 pm
by Spearthrower » Aug 15, 2023 5:04 pm
#973 ![]() by pfrankinstein » Aug 15, 2023 6:13 pm
by pfrankinstein » Aug 15, 2023 6:13 pm
Spearthrower wrote:And this declaration is based upon what data? Survey/poll results link?
I wonder how they created a survey that not only delved into the philosophical reasoning for a person's rejection of theism, but also quantified their intelligence at the same time.
Also, false dilemma - it's entirely reasonable to be both agnostic (i.e. don't believe there's any way to gain knowledge of alleged gods) and atheist (not believe in gods).
#974 ![]() by romansh » Aug 15, 2023 7:22 pm
by romansh » Aug 15, 2023 7:22 pm
Bertrand Russell wrote:As a philosopher, if I were speaking to a purely philosophic audience I should say that I ought to describe myself as an Agnostic, because I do not think that there is a conclusive argument by which one can prove that there is not a God. On the other hand, if I am to convey the right impression to the ordinary man in the street I think that I ought to say that I am an Atheist, because, when I say that I cannot prove that there is not a God, I ought to add equally that I cannot prove that there are not the Homeric gods.

#975 ![]() by Fenrir » Aug 15, 2023 9:31 pm
by Fenrir » Aug 15, 2023 9:31 pm

")
#976 ![]() by THWOTH » Aug 15, 2023 9:34 pm
by THWOTH » Aug 15, 2023 9:34 pm
#977 ![]() by Spearthrower » Aug 16, 2023 3:29 am
by Spearthrower » Aug 16, 2023 3:29 am
pfrankinstein wrote:Spearthrower wrote:And this declaration is based upon what data? Survey/poll results link?
I wonder how they created a survey that not only delved into the philosophical reasoning for a person's rejection of theism, but also quantified their intelligence at the same time.
Also, false dilemma - it's entirely reasonable to be both agnostic (i.e. don't believe there's any way to gain knowledge of alleged gods) and atheist (not believe in gods).
Most intelligent atheists are really agnostic,; they just don't realise it.
With venom the athiest.
Paul.
Scientific skepticism or rational skepticism (also spelled scepticism), sometimes referred to as skeptical inquiry,[1] is a position in which one questions the veracity of claims lacking empirical evidence.
#978 ![]() by Radicalthought0000 » Aug 27, 2023 11:01 pm
by Radicalthought0000 » Aug 27, 2023 11:01 pm
#979 ![]() by Spearthrower » Aug 28, 2023 4:15 am
by Spearthrower » Aug 28, 2023 4:15 am
Radicalthought0000 wrote:Atheism irrational, no. It is just... incomplete, have any of you practiced "empirical" spirituality? You need to if you want to be totally sure that there isn't anything spiritual and all is matter, no?
#980 ![]() by Radicalthought0000 » Aug 28, 2023 5:27 am
by Radicalthought0000 » Aug 28, 2023 5:27 am
Go ahead and outline the practice of 'empirical spirituality', otherwise how can anyone know whether they've done it or not?
How does 'empirical spirituality' provide any certainty about the existence of spirituality without circularity?
What does the term 'empirical' mean of something other than matter?
Users viewing this topic: No registered users and 1 guest